Location: Home >> Detail
Crop Breed Genet Genom. 2024;6(2):e240002. https://doi.org/10.20900/cbgg20240002

Research Institute of Green Science and Technology, Shizuoka University, Shizuoka 422-8529, Japan
To create a lodging-resistant isogenic line, the semidwarf gene Jukkoku_sd1 was incorporated into Koshihikari through multiple continuous backcrosses. The genome of this isogenic line, which was substituted with that of Koshihikari except for the Jukkoku_sd1 gene, was fully analyzed using next-generation sequencing. This genomic analysis led to the development of a novel isogenic lodging-resistant cultivar, 'Koshihikari sd1'. RNA analysis confirmed the transcription of the Jukkoku-derived sd1 gene in 'Koshihikari sd1', marking the first evidence of sd1 transcription. Furthermore, the taste and quality of 'Koshihikari sd1' were found to be equivalent to that of Koshihikari. 'Koshihikari sd1' is registered as Hikarishinseiki under USDA Plant Variety Protection No. 201000072.
The introduction of high-yielding semidwarf wheat and rice varieties significantly boosted global cereal production, more than doubling it between 1960 and 1990 [1]. The semidwarf rice variety IR8, developed using the Chinese landrace 'Dee-geo-woo-gen' (DGWG) and released by the International Rice Research Institute (IRRI), was known as "miracle rice" for its ability to respond well to fertilizer and produce increased yields without elongation of the culm. This variety's widespread adoption sparked a global "green revolution", especially in Asia's monsoonal regions where typhoons are common during harvest [1,2].
The semidwarf trait is a critical agronomic characteristic in crop breeding. Various countries have developed short-culm cultivars using independent sources of semidwarfism, such as the Japanese indigenous landrace 'Jukkoku' [3] or γ-ray-induced semidwarf cultivars like 'Reimei' in Japan and 'Calrose 76' in the USA [4]. Dwarf rice varieties have globally improved and stabilized yields [5,6], and their short stature is usually controlled by a single gene, sd1 [7-10]. The sd1 allele, located on the long arm of chromosome 1 [11-13], confers the semidwarf phenotype without adversely affecting grain yield [14-18]. The sd1 gene is a defective GA20ox-2, which encodes a C20-oxidase in the gibberellin biosynthesis pathway (GA 20-oxidase, OsGA20ox2) [8-10].
In Japan, Koshihikari frequently suffers lodging damage due to powerful typhoons, making the development of lodging-resistant cultivars a longstanding challenge. Koshihikari accounts for 33% of Japan's rice cropping area [19] and is increasingly demanded in the US and other countries due to its high quality and the growing popularity of Japanese cuisine [20]. However, lodging during heavy rainstorms presents a significant problem. To address this, the author developed a dwarf Koshihikari-type rice variety, 'Koshihikari sd1', offering greater lodging resistance than Koshihikari while maintaining over 99.9% of the Koshihikari genome, and has been registered in Japan and the USA.
Semidwarf Isogenic by Jukkoku_sd1Development of 'Koshihikari sd1' was started at first crossing 'Jukkoku' (sd1sd1) with 'Kanto No. 79' (Sd1Sd1), an early-maturing mutant of 'Koshihikari' [21]. The pedigree method was conducted in breeding program during the later generations of 'Kanto 79' × 'Jukkoku', and then the dwarf sd1 homozygous line ('Jukkoku'-type 'Koshihikari') whose heading date was the same as 'Koshihikari', was selected and fixed in the F4 generation. The flanking substituted region adjacent to sd1 had been restricted by fourteen recurrent backcrossing between the Sd1sd1 descendants of 'Jikkoku'-type 'Koshihikari' short line and 'Koshihikari' (Sd1Sd1) as the recurrent parent. The semidwarf phenotype (sd1sd1) was done by the BC14F3 generation, therefore in which 'Koshihikari sd1' line carried ≥99.9% of 'Koshihikari' background in their genome.
Whole genome sequencing was performed on both Koshihikari and 'Koshihikari sd1' [22]. Leaves were ground using a mortar and pestle while frozen in liquid nitrogen. Genomic DNA was extracted using the CTAB method, fragmented, and tagged to achieve approximately 500 bp fragments using the Nextera® transposome (Illumina Inc., San Diego, CA). After purification, adaptor sequences, including sequencing primers, were synthesized at both ends of each DNA fragment via a limited-time polymerase chain reaction, followed by size selection using AMPure XP magnetic beads (Beckman Coulter, Brea, CA). Sequencing data were obtained with paired-end 2 × 100 bp reads on a HiSeq 2500 next-gen sequencer (Illumina Inc., San Diego, CA). The raw reads were quality checked by FastQC (version 0.11.9; Babraham Institute), and low-quality sequences were trimmed using Trimmomatic (version 0.39) [23]. Reads from Koshihikari were mapped to the Nipponbare genome reference using Burrows-Wheeler Aligner (BWA) software (version bwa-0.7.17.tar.bz2) [24], and duplicates were removed using Picard (version 2.25.5) (http://broadinstitute.github.io/picard) and SAMtools (version 1.10.2) [25]. The consensus sequence of the Koshihikari genome was then constructed. Next, the read sequences from 'Koshihikari sd1' were mapped using the Koshihikari consensus genome as a reference. Single nucleotide variant (SNV) detection and SNV matrix generation were performed using GATK version 4.1.7.0 [26].
Using next-generation sequencing, 66,093,171 reads with an average length of 124 bp were obtained from Koshihikari and 1,267,339,019 reads with an average length of 125 bp from 'Koshihikari sd1'. By mapping 99.91% of the Koshihikari reads using the Nipponbare genome as a reference, a Koshihikari consensus sequence of 374,308,257 bp with a mean coverage of 35.68 was obtained. Similarly, 99.88% of the 'Koshihikari sd1' reads were assigned to the Koshihikari consensus sequence with a mean coverage of 22.42. A VCF file of 'Koshihikari sd1' was then constructed by SNP calling against the Koshihikari consensus sequence. Figure 1 displays the frequency distribution of single nucleotide polymorphisms on chromosome 1 of 'Koshihikari sd1' compared to Koshihikari. Aside from the region around 38 Mb from the 5' end of chromosome 1, SNPs were fewer than 10 per 0.1 Mb. This indicates that most of 'Koshihikari sd1' was substituted into the 'Koshihikari' genome after 14 backcrosses targeting the semidwarfing gene sd1. Jukkoku-derived SNPs associated with sd1 were identified near 38 Mb on the long arm of chromosome 1 (Figure 1). A single SNP from G to T was detected at 38,267,510 bp in the Sd1/sd1 locus (Os01t0883800-01: 38,267,149-38,270,233 bp) at the distal end of the short arm of chromosome 1 in the 'Koshihikari sd1' genome (Figure 1). Thus, the semidwarf rice 'Koshihikari sd1' has a nearly homogeneous Koshihikari genome except for the sd1 region, as confirmed by next-generation sequencing.
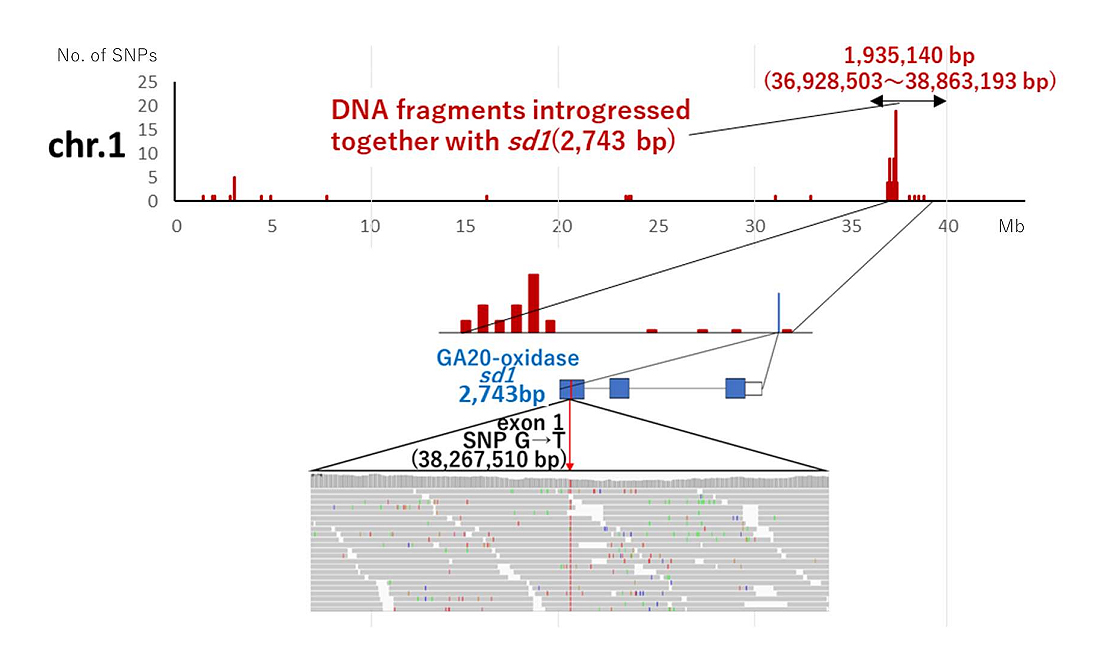 Figure 1. SNP-concentrated region derived from Jukkoku on chromosome1 surveyed by whole genome sequencing of 'Koshihikari sd1' (B14F3) against the genome of Koshihikari.
Figure 1. SNP-concentrated region derived from Jukkoku on chromosome1 surveyed by whole genome sequencing of 'Koshihikari sd1' (B14F3) against the genome of Koshihikari.
A single SNP from G to T was detected at 38,267,510 bp in the Sd1/sd1 locus (Os01t0883800-01: 38,267,149-38,270,233 bp) at the distal end of the short arm of chromosome 1 in the 'Koshihikari sd1' genome.
Jukkoku_sd1 Is Active AlleleThe sd1 allele on the long arm of chromosome 1, encoding a defective C20-oxidase in the gibberellin biosynthesis pathway (GA 20-oxidase, OsGA20ox2), confers the semidwarf phenotype that significantly contributed to the Green Revolution in rice. The dwarf characteristic of 'Koshihikari sd1' is derived from the sd1 gene. However, whether the sd1 gene in 'Koshihikari sd1' is transcribed remained uncertain. To investigate this, expression analysis was conducted using RT-PCR and PmaCI restriction enzyme digestion.
RNA extracted from the leaves and roots of 'Koshihikari sd1' and Koshihikari germinated in a Petri dish was used for RT-PCR. The reverse transcription reaction was performed with an RT primer (R6: TCAGCTGGCCGCCTCGACCTGCGCCG) designed from the 3' end of sd1 (Os01g0883800). RT-PCR was conducted with 24 primer combinations: four upstream primers (F1: GGAGCCCAAGATCCCGGAGCCATTCGTG, F2: CGACCTGAGGATGGAGCCCAAGATCCCG, F3: GACTCCACCGCCGGCTCTGGCATTGC, F4: CACGCCACCACAGCCGCACCAACCAC) from the first exon of sd1 and six downstream primers (R1: GAGGGTGCTGGAGAAGTAGTCGGCGAC, R2: CACCCTCCCCATTGGCGCGAAGTCGG, R3: TACCATGAAGGTGTCGCCGATGTTGATGACC, R4: CGTTCGACAGCGCCATGAAGGTGTCGCC, R5: TCAGCTGGCCGCCTCGACCTG, and R6: TCAGCTGGCCGCCTCGACCTGCGCCG) from the 3' end. A 779 bp fragment was amplified using primer F1 from the first exon and primer R4 from the second to third exon range, containing only exons. Additional fragments of 1316, 1304, 1400, and 1366 bp were amplified using upstream primers F1~F4 and downstream primer R6, containing the second intron. Fragments of 1428, 1416, and 1478 bp were obtained using upstream primers F1~F3 and RT primer R5, containing both the first and second introns (Figure 2). Primer design was based on the public sequence of Nipponbare (http://rgp.dna.affrc.go.jp). Since sd1 confers GA20ox-2 and shares homology with GA20ox-1 (Os03g0856700), primers were designed from regions with low homology between GA20ox-2 and GA20ox-1, ensuring distinct labeling of GA20ox-2 and GA20ox-1. PCR amplification was performed in a 20 μL reaction containing 10 ng genomic DNA, 1 μM of each primer, 0.4 mM dNTPs, 1 × GC buffer I, 2.5 mM MgCl2, and 0.5 units of LA Taq polymerase (Takara), with cycling conditions: 35 cycles of 94 ºC for 30 s, 58 ºC for 30 s, and 72 ºC for 1 min. PCR products were treated with PmaCI (CAC↓GTG), which detects a single nucleotide substitution site in the first exon of sd1.
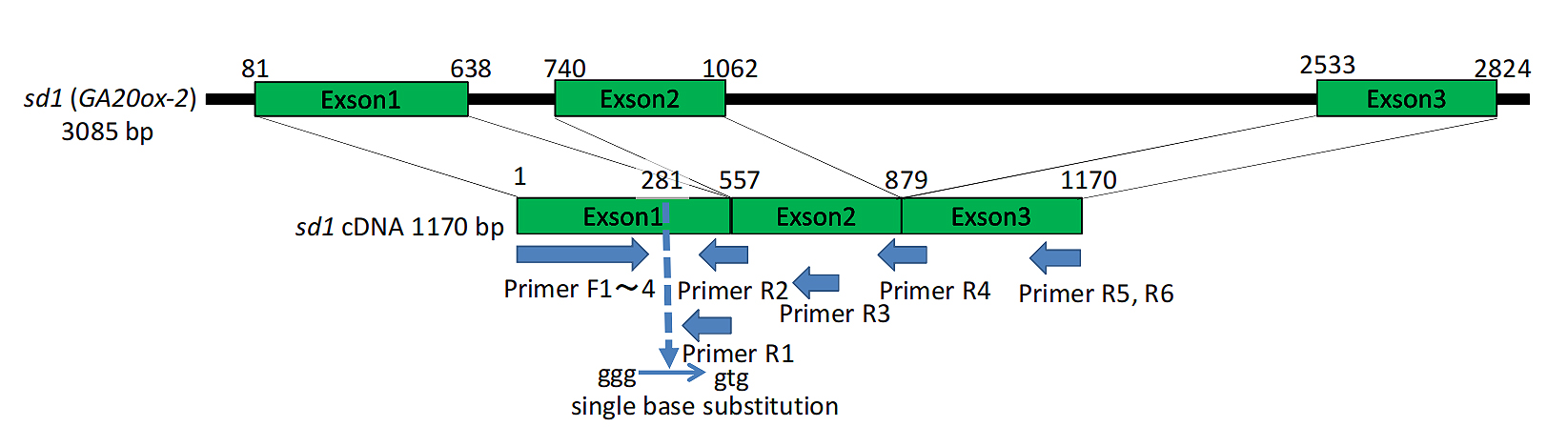 Figure 2. PCR primers designed for amplification of cDNA derived from sd1 (GA20ox-2) transcript.
Figure 2. PCR primers designed for amplification of cDNA derived from sd1 (GA20ox-2) transcript.
The reverse transcription reaction was performed with an RT primer (R6: TCAGCTGGCCGCCTCGACCTGCGCCG) designed from the 3' end of sd1 (Os01g0883800). RT-PCR was conducted with 24 primer combinations: four upstream primers (F1: GGAGCCCAAGATCCCGGAGCCATTCGTG, F2: CGACCTGAGGATGGAGCCCAAGATCCCG, F3: GACTCCACCGCCGGCTCTGGCATTGC, F4: CACGCCACCACAGCCGCACCAACCAC) from the first exon of sd1 and six downstream primers (R1: GAGGGTGCTGGAGAAGTAGTCGGCGAC, R2: CACCCTCCCCATTGGCGCGAAGTCGG, R3: TACCATGAAGGTGTCGCCGATGTTGATGACC, R4: CGTTCGACAGCGCCATGAAGGTGTCGCC, R5: TCAGCTGGCCGCCTCGACCTG, and R6: TCAGCTGGCCGCCTCGACCTGCGCCG) from the 3' end. The target fragment was detected with eight primer combinations. RNA extracted from the leaves and roots was used as the template, and the 779 bp fragment amplified with primers F1 and R4 was cleaved into 613 bp and 166 bp fragments by PmaCI digestion in 'Koshihikari sd1', but not in Koshihikari (Figure 3). Therefore, the sd1 gene from Jukkoku was confirmed to be transcribed in 'Koshihikari sd1'. This study provides the first evidence of the transcription of sd1, a defective gene of GA20ox-2.
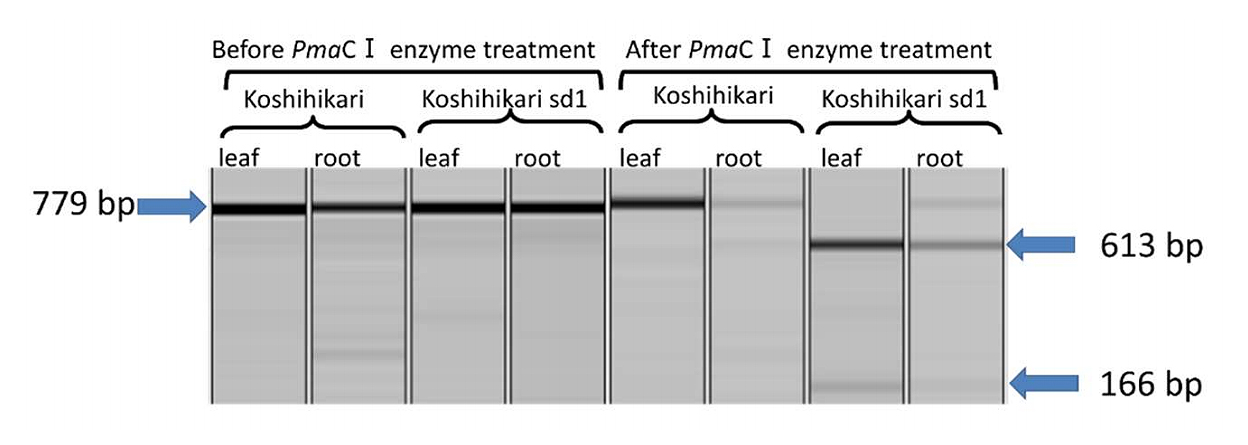 Figure 3. PCR amplification of cDNA from sd1 (GA20ox-2) transcript and their PmaCI-digestion.
Figure 3. PCR amplification of cDNA from sd1 (GA20ox-2) transcript and their PmaCI-digestion.
Left four lanes are PCR product by primers F1 and R4, and right four lanes show the fragments after PmaCI-digestion. Template RNAs ware extracted from leaf and root of Koshihikari and 'Koshihikari sd1', respectively. PmaCI split the PCR product from Jukkoku_sd1 in 'Koshihikari sd1' into two fragments, while the PCR product from Sd1 in Kosihikari remained as a single fragment.
Yield Performance of Jukkoku_sd1 in the Isogenic BackgroundPerformance tests were conducted in 13 experimental fields across Japan, including Miyagi, Ibaraki, Kanagawa, Nagano, Mie, Toyama, Ishikawa, Kyoto, Wakayama, Hyogo, Shimane, Okayama, Tokushima, Ehime, Oita, and Kumamoto (Table 1). 'Koshihikari sd1' and 'Koshihikari' were sown, and 128 seedlings per plot were transplanted with two replications. The heading date was recorded once 50% of all panicles had emerged from the flag leaf sheath. Days-to-heading was calculated from sowing date to heading date. Measurements of culm length, panicle length, number of panicles, leaf length, and leaf width were taken from 10 typical plants per genotype in each plot. Thousand-grain weight, grain yield of brown rice, grain quality, and eating quality were measured on bulks of 50 individuals. The means of each trait of the developed isogenic lines were statistically evaluated using the t-test to determine significant differences from Koshihikari.
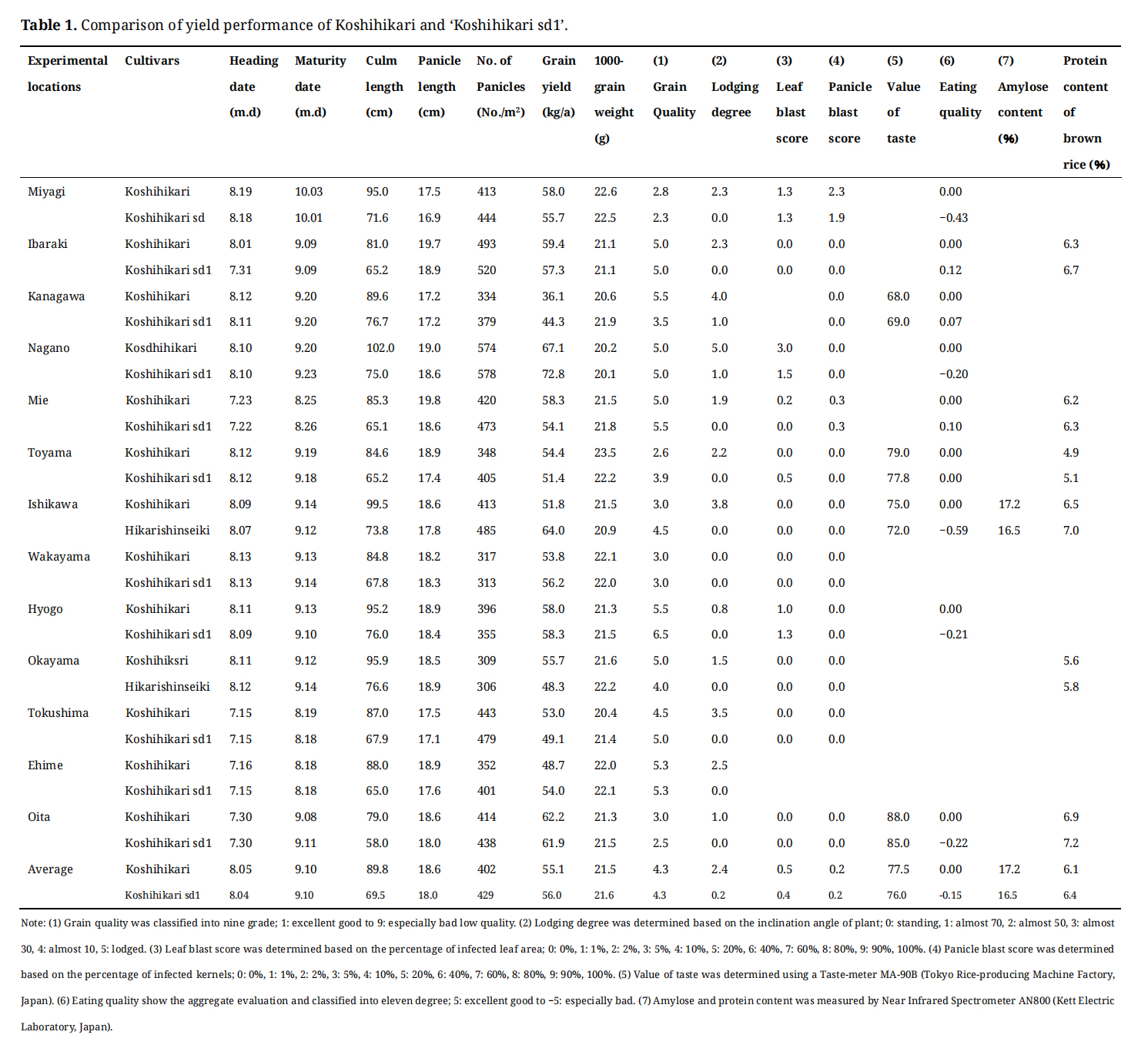 Table 1. Comparison of yield performance of Koshihikari and 'Koshihikari sd1'.
Table 1. Comparison of yield performance of Koshihikari and 'Koshihikari sd1'.
Heading: 'Koshihikari sd1' headed on average 0.25 days earlier than Koshihikari.
Plant height: The average height of 'Koshihikari sd1' is 69.5 cm (77% of Koshihikari), which is 20.3 cm (23%) shorter than Koshihikari on average (Figure 4).
 Figure 4. Phenotypic alteration from Kishishikri (left) to Koshishikri sd1 (right).
Figure 4. Phenotypic alteration from Kishishikri (left) to Koshishikri sd1 (right).
The average height of 'Koshihikari sd1' is 69.5 cm (77% of Koshihikari), which is 20.3 cm (23%) shorter than Koshihikari on average.
Lodging resistance: The average degree of lodging was 0.2, indicating significant improvement in lodging resistance.
Panicle length: Panicle length was 18.0 cm (97% that of Koshihikari).
Number of panicles: The average number of panicles was 429/m2 (107% that of Koshihikari). In Ishikawa prefecture, the number of panicles was 10% higher than Koshihikari.
Brown rice yield: The average yield was 56.0 kg/a (102% that of Koshihikari). In some regions, the yield was ≥7% higher than Koshihikari (Kanagawa: +23%, Nagano: +8%, Ishikawa: +24%, Ehime: +13%), while in other areas (Toyama and Tokushima) it was ≤5% lower, indicating regional differences in yield.
Grain weight of brown rice: The average grain weight of 'Koshihikari sd1' brown rice was 21.6 g (100% that of Koshihikari).
Brown rice quality: The average quality score was 4.3, the same as Koshihikari, indicating 'medium' quality.
Taste: The overall taste score was -0.15, indicating 'better than average' taste, equivalent to Koshihikari.
In summary, 'Koshihikari sd1' is a promising alternative to Koshihikari, offering resistance to typhoon damage, ease of cultivation, and comparable taste and quality across Japan. It is registered as Hikarishinseiki under USDA Plant Variety Protection No. 201000072 and certified as a brand rice in multiple Japanese prefectures of Okayama, Tottori, Tokushima, Niigata, Kochi, Shiga, Mie, Kagawa, Hyogo, Kyoto, Hiroshima, Tochigi, Kumamoto, Chiba, and Wakayama by the Ministry of Agriculture, Forestry and Fisheries of Japan.
All data generated from the study are available in the manuscript.
The author declares that they have no conflicts of interest.
The author would like to express gratitude to the Japan Science and Technology Agency (JST) for the Grant-in-Aid for Adaptable and Seamless Technology Transfer Program through Target-driven R&D (No. 08150094 and No. 08001167) that supported this work.
Special thanks to all those who provided test data from various prefectures across Japan.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
Tomita M. Genomics-Driven Development of Jukkoku_sd1-introgressed Isogenic Rice Koshihikari USDA 201000072. Crop Breed Genet Genom. 2024;6(2):e240002. https://doi.org/10.20900/cbgg20240002

Copyright © 2024 Hapres Co., Ltd. Privacy Policy | Terms and Conditions




