Location: Home >> Detail
Crop Breed Genet Genom. 2021;3(1):e210003. https://doi.org/10.20900/cbgg20210003
 ,
Chinedu Emmanuel Eze 2,
Diakaridia Traore 3,
Abebe Menkir 4
,
Chinedu Emmanuel Eze 2,
Diakaridia Traore 3,
Abebe Menkir 4
1 Department of Crop Production and Protection, Obafemi Awolowo University, Ile-Ife, Osun State 220005, Nigeria
2 Department of Agronomy, Michael Okpara University of Agriculture Umudike, Abia State 440109, Nigeria
3 Département de Biologie, Université des Sciences, des Techniques et des Technologies, de Bamako, 123456, Mali
4 International Institute of Tropical Agriculture, Ibadan 200001, Nigeria
* Correspondence: Richard Olutayo Akinwale.
This article belongs to the Virtual Special Issue "Genetic Gains in Plant Breeding"
Proper understanding of the mode of gene action in open-pollinated varieties (OPVs) maize parents helps breeder in the choice of appropriate breeding method to improve its genetic value. The objectives of the present study were to determine gene action controlling grain yield and other agronomic traits of late maturing elite OPVs and classify the varieties into heterotic groups. Ninety-one hybrids generated from 14 elite OPV parents using diallel mating design were evaluated with their parents plus three commercial checks under marginal rainfall, drought, and optimal environments in Nigeria from 2017 to 2018. The experiment was laid out in a 9 × 12 alpha lattice with three replications in each environment. Results showed that there were significant mean squares for grain yield and most agronomic traits. Significant general combining ability (GCA) and specific combining ability (SCA) mean squares for some of the traits indicated that additive and non-additive genetic gene actions were important in the inheritance of yield and those traits among this set of genotypes. However, non-additive genetic effects were more important than additive for grain yield and other agronomic traits in all research environments. Only TZL Comp-3 C3 DT had significant and positive GCA effects (0.336*) while three hybrids had significant and positive SCA for grain yield across research environments. Because of the preponderance of non-additive gene action over additive gene action, distinct heterotic groups could not be identified but four tester groups were identified by HSGCA (Heterotic grouping based on Specific and General Combining Ability) and three groups by HGCAMT (Heterotic grouping based on the GCA of Multiple Traits). Results of grouping were not related to the endosperm colour but grouping based on HGCAMT was related to the pedigree of the varieties. In conclusion, this study has demonstrated preponderance of non-additive gene action over the additive gene action for all measured traits. The presence of the non-additive gene action present in the studied materials can enhance identification of outstanding varietal hybrids and population testers that can serve as base genetic materials for future maize improvement through reciprocal recurrent selection program in SSA.
Maize is a major food security crop that supports millions of people in sub-Saharan Africa (SSA) and the rest of the developing world. When compared to developed countries, the low maize yield in SSA (1.5–2.0 t•ha−1) is principally attributed to production constraints, comprising of several abiotic stress factors and low adaptation of exotic germplasm to target environments in the major maize production agro-ecological regions of the SSA [1,2]. Several maize breeding programs use open-pollinated varieties (OPVs) as a source of genetic variability and favourable alleles of target traits. Though there might be cases of low yield, OPVs may be economically viable and sustainable, reveal phenotypic plasticity, and present wide adaptability to growing environments [3]. The conventional maize hybrid seed is relatively too expensive for a smallholder rural farmer in SSA, and requires more input for its production. Thus, considering elite OPVs as interim solution to boost maize productivity in rural areas of SSA is justifiable. Semagn et al. [4] pointed out that OPVs continue to occupy a large proportion of the maize production area in SSA because they represent the quickest and the easiest way to get improved genetic materials to resource-poor farmers at an affordable price.
Improvement in yield and yield components of OPVs may be achieved through crosses, which enhances the exploitation of the intervarietal heterotic effects, allelic complementarity, as well as gene actions and effects [5]. Even though farmers in this region are now been discouraged from cultivating OPVs because of their poor yield, studies have shown that hybrids formed from them exhibit outstanding heterotic potential, in terms of productivity across stressed and non-stressed environments when the heterotic patterns among the varieties are exploited [6–8].
Breeders make thousands of crosses and evaluate grain yield in resulting F1 plants in replicated field experiments of lines from known or unknown sources. By classifying these lines into heterotic groups, the development and evaluation of crosses that should be discarded would be avoided, thereby, allowing the exploitation of maximum heterosis by crossing lines belonging to different heterotic clusters [9]. Lines extracted from complementary populations developed from the parents of the opposite heterotic clusters usually show high combining ability with lines from the complementary population. Thus, the success of a hybrid program partly depends on the availability of information regarding the heterotic groupings of the parental lines. According to Badu-Apraku and Fakorede [10], grouping of maize inbred lines into appropriate heterotic groups defines the potential usefulness of such inbreds in a breeding program since it allows a proper understanding of the genetic relationships that exist within the inbred lines and enhances their efficient utilization in the development of hybrids, synthetic varieties, and subsequent formation of heterotic populations.
Several researchers have used the SCA effects of grain yield to classify maize genotypes into heterotic groups [11–14]. However, SCA effects for grain yield have been found to be influenced often by the interaction between two maize genotypes and the environment. Most times, this has led to the classification of the same genotypes into different heterotic groups in different studies [15,16]. Fan et al. [15] proposed the use of heterotic grouping based on Specific and General Combining Ability (HSGCA) method by combining both SCA and GCA effects of grain yield as a more appropriate method for assigning genotypes into heterotic groups. However, Badu-Apraku et al. [16] indicated that heterotic grouping of inbred lines based on one trait (grain yield) poses a challenge due to the complexity of this trait. Grain yield is controlled by several genes (polygenic), influenced by other traits and has low heritability under stress conditions. Bolaños and Edmeades [17] reported that selection for grain yield under drought conditions is inefficient due to the decline in the estimate of heritability of grain yield under environmental stress. Hence, heterotic grouping based on the GCA of multiple traits (HGCAMT) was proposed [16]. This method is based on measured multiple traits that are related to grain yield of genotypes with significant GCA effects across environments. Classification based on the GCA effects of multiple traits should be a better, realistic and more predictable approach for heterotic grouping of genotypes since GCA measures the additive gene effects of each trait. Badu-Apraku et al. [16,18] have successfully used this method to group early and extra-early inbred lines into heterotic clusters.
There has been a research gap in the improvement and utilization of elite OP maize varieties as an alternative and sustainable source of maize seed for smallholder farmers in SSA. With the dynamism associated with farmers’ environment, worsened by climate change, the availability of maize varieties with stable yields across contrasting environments would be a welcome development in this part of the world.
Hence, the objectives of the present study were to determine gene action controlling grain yield and other agronomic traits in late maturing elite OPVs and use the two heterotic grouping methods (HSGCA and HGCAMT) to classify the varieties into heterotic groups under rainforest agro-ecological conditions of Nigeria.
Field trials were carried out in 2017 and 2018 at the Teaching and Research Farms of Obafemi Awolowo University (OAU), Ile-Ife (7°31′ N, 4°31′ E, 256 m asl, and 1000–1250 mm annual rainfall) and Michael Okpara University of Agriculture, Umudike (05°29′ N, 07°33' E; 122 m asl, and 2177 mm annual rainfall) in Nigeria.
Generation of Diallel CrossesFourteen late/intermediate maturing elite OPVs of maize were selected from the drought-tolerant and pro-vitamin A breeding populations developed at the International Institute of Tropical Agriculture (IITA), Ibadan, Nigeria (Table 1). Each parental variety was planted into a 10-row plot of 5 m in length. The plants were nurtured under standard management practices. Ninety-one (91) variety hybrids were generated by making all possible crosses without reciprocals among the 14 elite OPVs following Griffing’s method 2 approach of diallel mating design. However, due to non-viability of the seeds of the parental varieties at evaluation, Griffing’s method 4 was used for the genetic analysis. The characteristics of the 14 OPVs involved in the diallel cross and the check varieties used are listed in Table 1. The crosses were developed using controlled hand pollination at the Teaching and Research Farm, OAU, Ile-Ife in 2017.
 Table 1. Characteristics of parents used for the diallel crosses and the check varieties.
Table 1. Characteristics of parents used for the diallel crosses and the check varieties.
The 14 parental varieties, their 91variety hybrids plus 3 commercial checks were evaluated in two locations under each of marginal rainfall, drought and optimal growing conditions in 2018 (Table 2). The checks were made up of 2 improved OPVs sourced from IITA and a local variety commonly grown by small-scale farmers in the locations. The growing conditions, which constituted six environments, were based on the time of planting and the total amount of rainfall. Under the marginal rainfall condition, the trials were set out at the onset of rainfall when the rain frequency is erratic and soil moisture is sub-optimal for maize cultivation. Under the optimal growing condition, the trials were established during the main planting season of maize with optimum amount of rainfall. Drought was achieved by planting towards the end of the rainy season (precisely third week in September), when flowering of the genetic materials was targeted to coincide with drought spell. The National Root Crops Research Institute agrometeorological unit (https://nrcri.gov.ng/index.php/agro-meteorology/) provided meteorological data for the location at Umudike, while the Micrometeorology Unit, Department of Physics, OAU provided that of the Ile-Ife location, which has its weather station at the experimental site. The experiment was laid out in a randomized incomplete block design (9 × 12 alpha lattice) with 3 replications in each environment. Experimental units consisted of 5 m two-row plots, with a spacing of 0.75 m. The intra row spacing was 0.50 m in all trials. Three seeds were planted, and the seedlings were later thinned down to two per hill at about 2 weeks after emergence to have a final plant population density of about 53,333 plants ha−1. A common 2-row border was used at the beginning and end of each block to remove bias that may be created as a result of undue advantage to entries that fell at the borders or extremes of the blocks.
 Table 2. Characteristics of the 6 test environments used for the evaluation of the genotypes.
Table 2. Characteristics of the 6 test environments used for the evaluation of the genotypes.
Data were recorded under all the growing conditions on days to 50% silking (DS) and days to anthesis (DA) as the number of days from planting to when 50% of plants in a plot had emerged silks and had shed pollen, respectively. The anthesis-silking interval (ASI) was calculated as the difference between DS and DA. Ear (EHT) and plant heights (PHT) were measured as the distance from the base of the plant to the height of the node bearing the upper ear and the first tassel branch, respectively. Stalk lodging (the percentage of plants broken at or below the highest ear node) and root lodging (the percentage of plants leaning more than 30° from the vertical) were also recorded. Plant aspect (PASP) was an assessment of overall plant architecture and appeal (plant and ear heights, uniformity of plant height, cob size, disease and insect damage and lodging) and was recorded on a scale of 1 to 5 where 1 = excellent plant type and 5 = poor plant type. Ear aspect (EASP) was based on freedom from disease and insect damage, ear size, uniformity of ears and will be recorded on a scale of 1 to 5, where 1 = clean, uniform, large, and well-filled ears and 5 = rotten, variable, small, and partially or poorly filled ears. Ear number per plant (EPP) was estimated by dividing the total number of ears harvested in a plot by the number of plants in that plot. Grain yield was estimated from the ear weight and converted to kg•ha−1. A shelling percentage of 80% was assumed for all genotypes and the grain yield was adjusted to 15% moisture [18] using the formula:

where γ = grain yield (kg•ha−1), ϵ = ear weight (kg•m−2), n = moisture at harvest, φ = plot area (m2), 85 = 100 − 15 (a factor for 15% moisture content adjustment).
Statistical AnalysisSeparate analysis of variances (ANOVAs) were performed on the data collected across locations for each research condition (marginal, drought and optimal growing) with PROC GLM in SAS using a RANDOM statement with the TEST option [19]. Subsequently, combined ANOVA was performed across the test environments for all the data collected. Environments, replicates and block were treated as random factors while entries were considered as fixed effects. The least significant difference (LSD) was used in separating means. The statistical model used for the ANOVA is as follows:

where Yijkg is the observed measurement for the gth genotype grown in the environment i, in the block k, in replicate j; u is the grand mean; Ei is the main effect of environment; Rj(i) is the effect of replicate nested within environment; Bk(ij) is the effect of block nested within replicate j by environment i; Gg is the effect of genotypes; GEgi is the interaction effect between genotype and environment, and εijkg the error term.
The combining ability analysis was carried out based on diallel Griffing’s method 4 approach The GCA and SCA effects of the parents and the crosses, respectively, as well as their mean squares in each and across the environments were estimated following Griffing’s method 4 model I [20]. Data for the parental varieties were excluded from the analysis due to poor and irregular germination of the seeds of the parental varieties during evaluation. The DIALLEL-SAS program developed by Zhang et al. [21] and adapted to SAS software version 9.4 was utilized. The t-test was used in testing both the GCA and SCA effects for significance. Their standard errors were estimated as the square root of the GCA and SCA variances [20].
The relative importance of GCA and SCA was investigated using both ratio of sum of squares of GCA to SCA and Baker’s approach [22] using the following formula:

where σ2gca = GCA variance components and σ2sca = SCA variance component. The closer the ratio is to one (1), the greater the predictability of a specific hybrid’s performance based on GCA alone. The proportions of the additive and non-additive gene actions were estimated from the GCA and SCA effects respectively. Similarly, proportion of GCA to SCA was computed on the ratio of sum of squares of GCA to the genotypic sum of squares by dividing sum of squares of GCA by genotypic sum of squares.
As proposed by Fan et al. [15], the HSGCA method was used to assign the OPVs into heterotic groups using GCA for grain yield of the parents and SCA for grain yield of their crosses, thus,

where Xij is the mean yield of the cross between the ith tester and the jth parent, Xi is the mean yield of the ith tester across jth parents. The HSGCA estimates were then subjected to Ward’s minimum variance cluster analysis using the JMP software, version 14 (SAS Institute Inc., Cary, NC, USA).
The statistical model used by the HGCAMT method to assign the parents into heterotic groups is represented as:

Where Y is HGCAMT, the genetic value measuring the relationship among genotypes based on the GCA of multiple traits i to n; Yi is the individual GCA effect of genotypes for trait i; Ȳi is the mean of GCA effects across genotypes for trait i; si is the standard deviation of the GCA effects of trait i; Єij is the residual of the model associated with the combination of parent i and trait j.
Heterotic grouping by the HGCAMT was carried out by standardizing the GCA effects (mean of zero and standard deviation of 1) of 10 considered traits. The standardization was done to minimize the effects of different scales of the traits. The standardized GCA effects were subsequently subjected to Ward’s minimum variance cluster analysis using the software JMP version 14 (SAS Institute Inc., Cary, NC, USA). Dendrograms were subsequently constructed for the groupings based on HSGCA and HGCAMT.
Results from the combined ANOVAs of the genotypes evaluated across the test environments indicated significant entry (G) and environment (E) effects for all traits except G for flowering traits and ear rot (Table 3). The entry mean squares, partitioned into its components, showed that GCA mean squares were significant for grain yield, emergence percentage (EMERG), DA and PHT while SCA was significant for all the traits except ear rot (EROT) across the test environments. The GCA × E interaction effect was shown to be significant for EASP, EMERG, ASI, and EROT. For all the traits measured across the test environments, no significant SCA × E interaction effects were recorded (Table 3).
The observed relative importance of GCA over SCA across the test environments, as given by the Baker ratio, was low for all the traits because the values are closer to zero than 1 (Table 3).
 Table 3. Mean squares and combining ability ratios for grain yield and other agronomic traits of 91 late maturing varietal hybrids of maize across test environments.
Table 3. Mean squares and combining ability ratios for grain yield and other agronomic traits of 91 late maturing varietal hybrids of maize across test environments.
The proportion of additive gene effects for GY was 27.6% across environments, 15.0% under marginal condition, 25.7% under drought condition and 26.3% under optimal condition. The lowest contribution of additive gene effects (10.3%) across the environments was recorded for ASI (Figure 1). The proportions of the non-additive genetic variance was greater than the additive genetic components in all environments for grain yield and other measured traits (Figure 1). Comparing the proportion of GCA to SCA under each study condition, the proportion of additive component of the total genetic variance was highest under drought conditions for EMERG, DA, DS, and ASI (Figure 1). Additive component was highest under marginal growing conditions for DA, PHT, and EROT while additive component was highest under optimal growing conditions for yield, PHT and EASP (Figure 1).
Positive and significant GCA effects for grain yield and emergence were observed for TZC3. However, its GCA effect for DA was negative and significant. Positive GCA effects were further observed for STRY2, TZLC1, DTSY14, DTSY2 and WDTS2 across environments with respect to grain yield (Table 4). The parents with negative GCA effects for PASP and EASP across the environments are STRY2, TZC3, WDTS2, and PVA2. Parents TZC3 and DTSY14 had significant and positive GCA effects for EMERG while PVA4 had a highly significant but negative GCA effect for EMERG. For DA, TZC3, TZLC1, and TZC4C2 had significant and negative GCA effects while STRY2 and PVA4 had significant but positive GCA effects. For PHT, all the parents had significant either positive or negative GCA effects except TZC3, DTSY14, PVA7 and WDTS1 whose effects were not significant (Table 4).
Three (3) crosses, IWD × TZC3 (0.885), TZLC1 × TZC4C2 (0.652), and STRY2 × TZC3 (0.647) had significant positive SCA effects for grain yield while STRY2 × WDTS1 had significant negative SCA effect for grain yield (Table not shown).
 Figure 1. Proportion of additive (lower blue bar) and non-additive (upper brown bar) genetic variance for grain yield (GY), plant aspect (PASP), ear aspect (EASP), ears per plant (EPP), emergence at 9 days after planting (EMERG), days to anthesis (DA), day to silking (DS), anthesis-silking interval (ASI), plant height (PHT), and ear rot (EROT) under drought, optimal and marginal growing conditions, and across environments. DT = drought condition, OPT= optimal growing condition, MAR = marginal condition, ACR= across environments.
Figure 1. Proportion of additive (lower blue bar) and non-additive (upper brown bar) genetic variance for grain yield (GY), plant aspect (PASP), ear aspect (EASP), ears per plant (EPP), emergence at 9 days after planting (EMERG), days to anthesis (DA), day to silking (DS), anthesis-silking interval (ASI), plant height (PHT), and ear rot (EROT) under drought, optimal and marginal growing conditions, and across environments. DT = drought condition, OPT= optimal growing condition, MAR = marginal condition, ACR= across environments.
 Table 4. General combining ability effects of open pollinated parents for grain yield and other traits across stress and non-stress environments.
Table 4. General combining ability effects of open pollinated parents for grain yield and other traits across stress and non-stress environments.
Since there was preponderance of SCA over GCA for grain yield and most traits under the study environments, it implies that distinct heterotic groups cannot be identified (Badu-Apraku et al, 2011). However, significant GCA and SCA mean squares from the results of the diallel crosses justify identification of tester groups among this set of parent. In the dendrograms constructed in Figures 2 and 3, tester groups were identified at R-square of approximately 0.4. Four tester groups were identified from the result of the cluster analysis of the grain yield data based on HSGCA across the test environments (Figure 2). The first group was made up of mostly drought tolerant OPVs consisting of white and yellow grains from various germplasm sources. The second group contained all the Pro Vitamin A (PVA) varieties except PVA2, which was classified into group three. IWD and WDTS1 made up group 4 which are white-grained and drought tolerant materials. Across the environments, three heterotic groups were identified based on HGCAMT (Figure 3). The OPVs, DTSY2 and DTSY14 constituted the first group, IWD and PVA4 belonged to the third group while the rest of the parental OPVs were classified into the second group. In clustering of the multiple traits, the grain yield was clustered along with EMERG and PHT while EASP, EROT and EPP formed a different cluster (Figure 3).
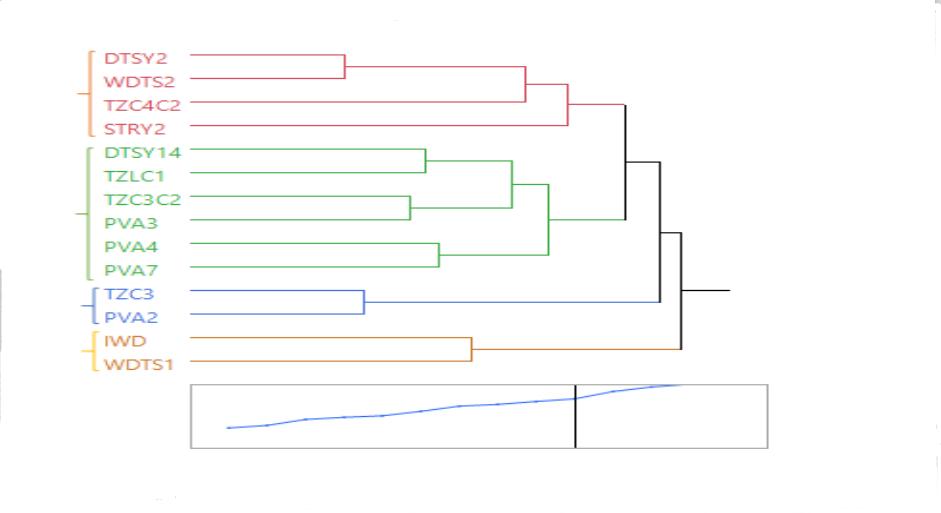 Figure 2. Dendrogram of 14 late maturing OP maize constructed from HSGCA effects method using Ward’s minimum variance cluster analysis across test environments. The number of clusters were determined at R2 = 0.4.
Figure 2. Dendrogram of 14 late maturing OP maize constructed from HSGCA effects method using Ward’s minimum variance cluster analysis across test environments. The number of clusters were determined at R2 = 0.4.
 Figure 3. Dendrogram of 14 late maturing OP maize constructed from GCA effects of grain yield and other traits (HGCAMT) using Ward’s minimum variance cluster analysis across test environments. The number of clusters were determined at R2 = 0.4.
Figure 3. Dendrogram of 14 late maturing OP maize constructed from GCA effects of grain yield and other traits (HGCAMT) using Ward’s minimum variance cluster analysis across test environments. The number of clusters were determined at R2 = 0.4.
In summary, the tester groups by the different grouping methods are presented in Table 5. DTSY2 and DTSY14 were grouped together by HGCAMT; the case was different with HSGCA method. In addition, the HGCAMT method grouped WDTS1 and WDTS2 together across the test environments. However, they were grouped in different clusters by HSGCA method.
 Table 5. Summary of the heterotic groups of 14 late-maturing OPV maize identified by different heterotic grouping methods across the test environments.
Table 5. Summary of the heterotic groups of 14 late-maturing OPV maize identified by different heterotic grouping methods across the test environments.
Correlation among traits measured revealed that grain yield had highly significant correlations with plant aspect (r = −0.62 **), ear aspect (r = −0.51 **), ears per plant (r = 0.53 **), and plant height (r = −0.73 **). Similarly, yield was also significantly correlated (p < 0.05) with days to anthesis (r = −0.36 *), days to silk (r = −0.43 **), and number of ear rot (r = 0.47 **) but the strength of correlation coefficients were weaker (Table 6).
 Table 6. Correlation matrix between grain yield (GY) and other agronomic traits considered across environments.
Table 6. Correlation matrix between grain yield (GY) and other agronomic traits considered across environments.
The presence of significant means squares for grain yield and some agronomic traits indicated that there was substantial genetic variability among the genotypes upon which genetic improvement program can be based. When genotypic variability is partitioned into its components, the result revealed significant GCA and SCA mean squares for most important traits under each and across all test environments indicating that both additive and non-additive gene actions are important in the inheritance of most traits among the set of genotypes been studied. In addition, the result also implies that wide scope for improvement exists for some of the measured traits using hybridization, recurrent selection and backcrossing methods to develop varietal hybrids, synthetics as well as population.
It was observed that both additive and non-additive gene actions were involved in the inheritance of all the considered traits. However, there was the preponderance of SCA over GCA mean squares for grain yield and other traits measured in all the contrasting environments with the relative importance of GCA to SCA effects for grain yield and ear aspect increasing from stress to non-stress environments while most of the other traits decreased from stress to non-stress environments. In the same vein, the low Baker’s ratios also indicate the predominance of non-additive gene action over additive gene action; hence, a low predictability of progenies performance from parents’ GCA effects. The performance of progeny in this set of crosses was better in specific combinations and therefore could not be predicted for a wide range of crosses. The result obtained suggests that non-additive gene action prevailed over the additive counterpart for these traits [18] and that the major component accounting for the differences among the OPVs evaluated was the SCA. The result is in agreement with the report of Adewale et al. [23] and Badu-Apraku et al. [18] who also reported preponderance of non-additive gene action over the additive gene action among early maturing maize inbred lines.
The present results are also in agreement with the findings of other researchers [24–27] who reported the preponderance of non-additive gene action for the expression and inheritance of yield and other traits measured in maize. The results of this study is in partial agreement with the findings of Wegary et al. [28] who reported SCA effects to be more important under optimal conditions and GCA effects, more important under drought for grain yield. The variations in the results reported in the two studies may be attributed to the different backgrounds of the materials used and might have contained some genes with different modes of action [16].
This current research suggested that under marginal, drought, and optimal growing conditions, the gene actions controlling grain yield and other measured traits were similar in this set of elite OPVs. In the work of Badu-Apraku et al. [18], similar gene actions (additive in this case) were responsible for controlling grain yield and most measured traits in the early maturing QPM inbred lines used. However, since additive gene action is significant, it can be deduced that progress can be made from selection for emergence percentage, days to anthesis, days to silking, and ASI under drought conditions because proportion of additive gene action was highest for these traits compared to the other traits. This result is in agreement with other studies where days to silking and ASI have been identified as drought adaptive traits [29–31]. Similarly, under marginal growing conditions, plant height, ear rot, and days to anthesis can be considered in a selection programme and under optimal growing conditions, yield, plant height and ear aspect can be selected for with good progress.
The results revealed a possibility to identify a potential tester under the contrasting environments which possess outstanding general combining abilities for desired traits and such tester can be used in subsequent studies for population improvement. The GCA effect of an open pollinated parent is a function of its relative importance as a parent for of population improvement and the development of synthetic varieties and as a tester for the improvement of a target trait in a population. Outstanding genotypes, in terms of GCA and SCA for grain yield and other agronomic traits, could be exploited for the development of heterotic populations that can serve as an initial gene pool for subsequent improvement and development of synthetic varieties and hybrids that are high yielding for the SSA region [32]. The positive and significant GCA effects (grain yield) observed for TZL Comp-3 C3 DT (TZC3) across research environments indicated that this variety possesses favourable alleles for grain yield and would contribute high yields to its progenies. Thus, it could serve as a tester for improving grain yield in a population. Similarly, the variety also possessed desirable GCA for emergence percentage and days to anthesis.
The combining ability estimate of a genotype across research environments is a measure of the performance and stability of that genotype in a hybrid combination or in a population development. Genotypes with outstanding GCA and SCA across research environments are suitable for hybrid and population development for the region of SSA [32]. Parent DTSY14 had significant negative GCA effects for ear aspect across the environments. This is an indication that this parent would contribute favourable alleles to ear aspect and indirectly to grain yield in their progenies since ear aspect is always closely associated with grain yield. Parents TZLC1, TZC3 and TZC4C2 showed significant and negative GCA effects for days to anthesis across research conditions. These results suggest that these OPVs will contribute favourable alleles to their progenies for earliness under contrasting environments. Mhike et al. [33] and Halilu et al. [34] reported similar significant GCA estimates for this trait and other agronomic traits.
The SCA effect of a particular cross determines its usefulness in exploiting heterosis and hybrid development. Across the test environments, only IWD × TZC3, TZLC1 × TZC4C2 and STRY2 × TZC3 showed significant positive SCA effects for grain yield. The cross, P4 × P6, involved two parents with positive GCA effects, suggesting an additive × additive type of gene action that can be fixed in subsequent generations if no repulsion phase linkages are involved [35]. On the other hand, IWD × TZC3 and TZLC1 × TZC4C2 involved crosses between contrasting parental lines indicating involvement of additive × dominant gene interactions in the two crosses. The high yield potential expressed by IWD × TZC3 and TZLC1 × TZC4C2 across test environments could be attributed to the contribution of favourable alleles from both poor and good combiners. This suggests the exploitation of heterosis in F1 generation as high yield potential might not easily be fixed in the subsequent generation. This finding is in line with the result of Meseka and Ishaaq [35] in maize, meanwhile, Peng and Virmani [36] had earlier reported the interaction between positive alleles from good combiners and negative alleles from poor combiners in rice hybrids.
As indicated by the significant GCA × environment interaction mean squares for some traits across test environments, the parents exhibited differential performance in hybrid combinations under the different environmental conditions in the study. Badu-Apraku et al. [18,37,38] reported similar results and this stresses the importance of testing OPVs in contrasting environments to identify those that have stable performance to be used in the development of stress tolerant synthetics and hybrids. It further underscores the earlier view of Kang [39] and Akinwale et al. [32] who stated that the environment plays prominent role in the phenotypic expression of agronomic characters, thereby suggesting that ignoring environmental component in the field would impede progress and advances in selection. The significant GCA × environment interaction provides vital information on the efficient use of an OPV parent as a tester in the different environments and suggests the need to select different parental lines for hybrid development under the research condition and at individual locations.
Identification of heterotic groups is fundamental in hybrid development because heterotic groups and patterns provide information for exploiting heterosis or hybrid vigour for the traits of interest. Important conditions for identifying distinct heterotic groups among a set of parent are significant additive and non-additive gene action and predominance of additive gene action over non-additive (Badu-Apraku et al. [38]. In this study, although there was significant mean squares for both GCA and SCA for most traits, the preponderance of non-additive gene action over additive gene action did not allow for identification of well-defined heterotic groups among the OPVs evaluated. However, tester groups were identified based on the dendrograms. Classification by the two methods in this study did not follow similar trends. The HSGCA and HGCAMT methods classified the 14 varieties into 4 and 3 groups respectively. Generally, it appeared classification based on HGCAMT identified clusters based more on pedigree and parentage as the two drought tolerant synthetic varieties were clustered together; three of the four provitamin A varieties were clustered into the same group; and all the TZL composite varieties were grouped together in cluster 2. On the contrary, considering the result of the analysis of variance and partitioning of the total genotypic variation into its components, the fact that the non-additive gene action was preponderant over the additive counterpart for grain yield and other traits; it is expedient to say that the grouping method that include SCA (non-additive effect) will be more efficient than the one that exclude the same effect in its classification. Thus, classification based on HGSCA is preferable in this study, which identified 4 groups. The grouping of the PVA varieties into separate clusters by the two methods indicates that there is wide variability among the provitamin A varieties and there is possibility of improving the agronomic performance of the provitamin A varieties.
In the grouping of early maturing QPM inbred lines using HSGCA and HGCAMT methods by Badu-Apraku et al. [18], HSGCA method also had one group more than HGCAMT method. The grouping was not related to the endosperm colour of the OPVs as the groups were consistently composed of OPVs from both endosperm colour types. Similar result was reported for 28 early maturing inbreds classified into heterotic groups based on combining ability by Akinwale et al. [32]. The classification of DTSY2 and DTSY14, WDTS1 and WDTS2 together in the same group across test environments by the HGCAMT method indicated that the grouping of the OPVs was based mainly on their pedigree and to a lesser degree on the reaction of the OPVs to the stress environments. This outcome is in line with the findings of several other authors [14,16,32].
Based on the analysis of relationship among traits, plant height, plant and ear aspects and ears per plant were identified as important secondary traits for indirect selection for grain yield across the studied conditions due to their significant correlation with it. This result is in agreement with findings of [40,41].
There was wide genetic variability among the varietal parents used for this study. Although, both additive and non-additive gene actions were significant in the control of grain yield and other agronomic traits, non-additive gene action was preponderant over additive gene action for all traits. The OPV, TZC3 (TZL Comp - 3 C3 DT) was identified as the best in terms of good general combining ability effects for grain yield and other traits across research environments. The favourable alleles from this parent should be harnessed for the development of high yielding and drought tolerant open pollinated varieties that can serve the rural maize farmers of the sub region in the face of climate change.
Four tester groups were identified among the 14 varieties from which population crosses could be made, which will serve as base population from where superior inbreds could be extracted.
Some of the dataset generated from this study are in the manuscript. The rest of the data is available from the authors upon request.
Richard Olutayo Akinwale conceptualized, laid out the field, supervised, collected data and guided in the data analysis and contribute to the writing of the paper.
Chinedu Emmanuel Eze carried out the fieldwork, field layout, data collection and analysis. He wrote the first draft of the article.
Diakaridia Traore contributed to the supervision of the research work.
Abebe Menkir supplied the genetic materials used and contributed to the methodology and carried out the editing of the manuscript before submission.
The authors declare no conflicting interest.
This work was supported financially by the German Federal Ministry of Education, through the West African Science Service Centre on Climate Change and Adapted Land-use (WASCAL) fellowship and an Ernst Mach Grant (Worldwide) from OeAD, the Austrian agency for International Mobility and Cooperation in Education, Science and Research for the second author.
The authors extend their gratitude to Dr. Baffour Badu-Apraku (IITA, Ibadan) for the provision of pollination bags used for the crosses. The authors also appreciate the contributions of Dr. Sebastian Michel and Professor Hermann Bürstmayr of the Department of Agrobiotechnology, Institute of Biotechnology in Plant Production Tulln, Austria, for their technical support in the latter phase of the work.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
Akinwale RO, Eze CE, Traore D, Menkir A. Detection of Non-Additive Gene Action within Elite Maize Populations Evaluated in Contrasting Environments under Rainforest Ecology in Nigeria. Crop Breed Genet Genom. 2021;3(1):e210003. https://doi.org/10.20900/cbgg20210003

Copyright © 2021 Hapres Co., Ltd. Privacy Policy | Terms and Conditions




