Location: Home >> Detail
Crop Breed Genet Genom. 2023;5(2):e230003. https://doi.org/10.20900/cbgg20230003

1 Department of Plant Sciences and Plant Pathology, Montana State University, Bozeman, MT 59717, USA
2 Western Agriculture Research Center, Montana State University, Corvallis, MT 59828, USA
* Correspondence: Andrej Svyantek.
This article belongs to the Virtual Special Issue "Eggplant Breeding and Genetics"
Eggplant (Solanum melongena L.), also known as aubergine or brinjal, is an economically important horticultural crop from the nightshade family (Solanaceae) which is cultivated for human consumption of its berry fruits. Numerous traits are important for eggplant productivity in diverse environments including disease resistance, abiotic stress resistance, and production traits. Breeding programs utilize heritability estimates to understand the proportional role of genetic factors in phenotypic expression of crops. Examining 60 published studies, this literature review summarizes ranges of observed broad-sense heritability in eggplants for ten traits related to plant architecture (plant height and branch number), phenology (days to first flower and days to fifty percent flowering), fruit characteristics, and production traits (fruit length, fruit girth, fruit diameter, individual fruit mass, and yield). Higher mean broad-sense heritability estimates were observed for yield components rather than yield per se. Due to the importance of population structure and environmental conditions on the estimate of heritability for quantitative traits, estimates are not transferrable; however, generally for all traits examined, breeding programs can expect targeted responses from selection.
BC, back cross; BNO, branch number; DFFL, days to first flower; D50F, days to fifty percent flowering; FRLN, fruit length; FRGR, fruit girth; FRDM, fruit diameter; FRNO, fruit number; FRMS, individual fruit mass; h2, broad-sense heritability; NCI, North Carolina Design I; NCII, North Carolina Design II; PLHT, plant height; YLD, yield
Eggplant (Solanum melongena L., 2n = 24) is the third most widely cultivated solanaceous vegetable crop after potatoes and tomatoes. Since FAOSTAT data collection began in 1961, the global production of eggplant has climbed gradually, reaching roughly 56 million tons in 2020 [1]. India and southern China are the two most commonly proposed places of origin since they have similarly ancient written evidence of eggplant use dating back about 2000 years [2]. China and India produce more than 80% of the world’s eggplant production [1,3,4]. There are approximately 750,000 total hectares dedicated to eggplant cultivation in both China and India (Figure 1). India exceeded 10,000,000 tons of eggplant produced in 2020 while China exceeded 30,000,000 tons of eggplants. Yield per unit of land has increased but it ranges widely from over 76 tons/ha to under 2 tons/ha across the ten nations which have the greatest cumulative eggplant production (Figure 2).
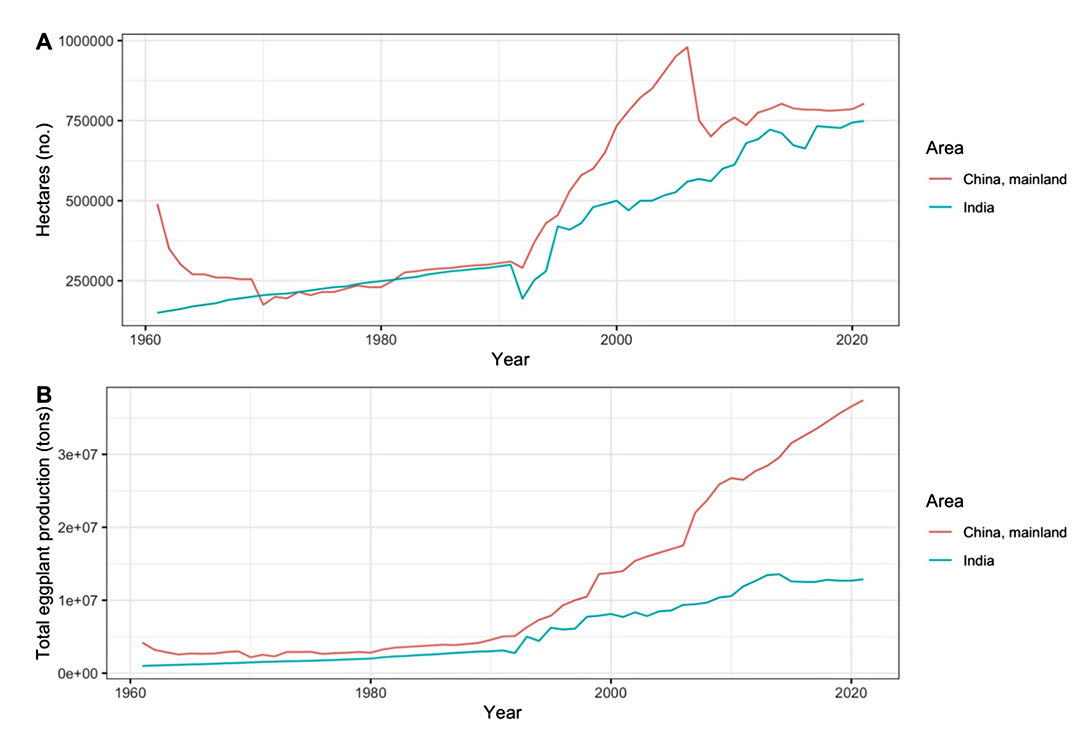 Figure 1. (A) Number of hectares dedicated to eggplant cultivation in China and India (1961–2021). (B) Total eggplant production in China and India (1961–2021).
Figure 1. (A) Number of hectares dedicated to eggplant cultivation in China and India (1961–2021). (B) Total eggplant production in China and India (1961–2021).
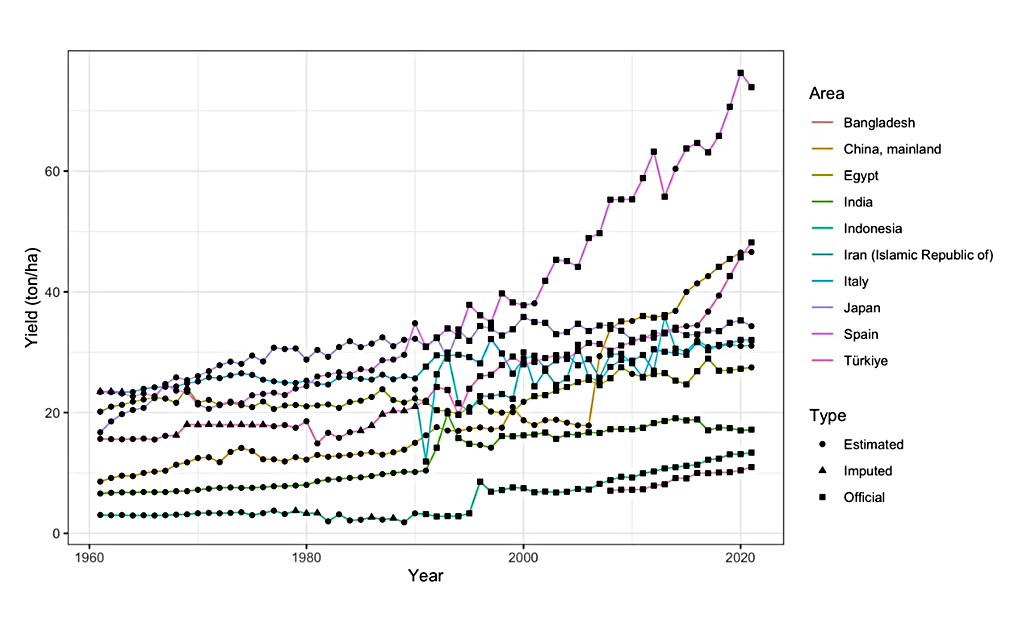 Figure 2. Average eggplant yield per hectare (1961–2021) for the ten nations with the greatest overall eggplant production in 2020.
Figure 2. Average eggplant yield per hectare (1961–2021) for the ten nations with the greatest overall eggplant production in 2020.
In the U.S., eggplants are predominately grown for fresh market, with 7564 operations with area harvested across 5365 acres based on the 2017 USDA Agriculture Census (Figure 3) [5]. The top five U.S. states based on acres harvested are New Jersey (n = 849), California (n = 705), Florida (n = 685), Georgia (n = 624), and New York (n = 264). The top five based on total number of operations with area harvested are California (n = 638), New York (n = 546), Pennsylvania (n = 449), New Jersey (n = 416) and Wisconsin (n = 332) (Figure 4). The price received by producers has increased from below 500 USD/ton in the late 1990s and early 2000s to over 1400 USD/ton in the early 2020s (Figure 5).
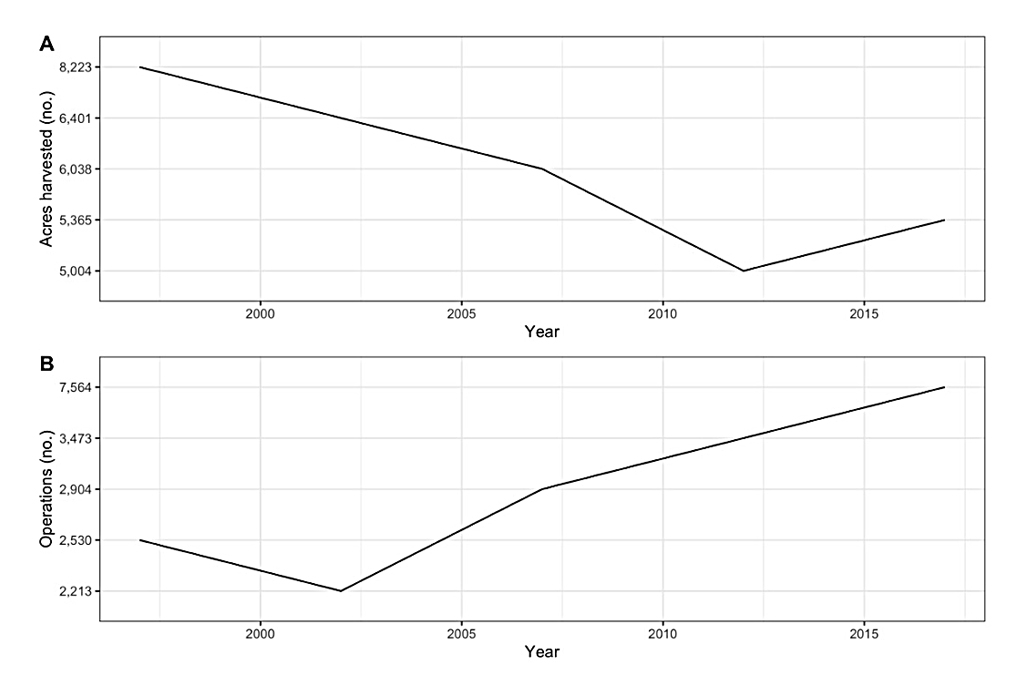 Figure 3. (A) Total acres of eggplants harvested in the U.S. (1997–2017). (B) Total number of operations producing eggplants in the U.S. (1997–2017).
Figure 3. (A) Total acres of eggplants harvested in the U.S. (1997–2017). (B) Total number of operations producing eggplants in the U.S. (1997–2017).
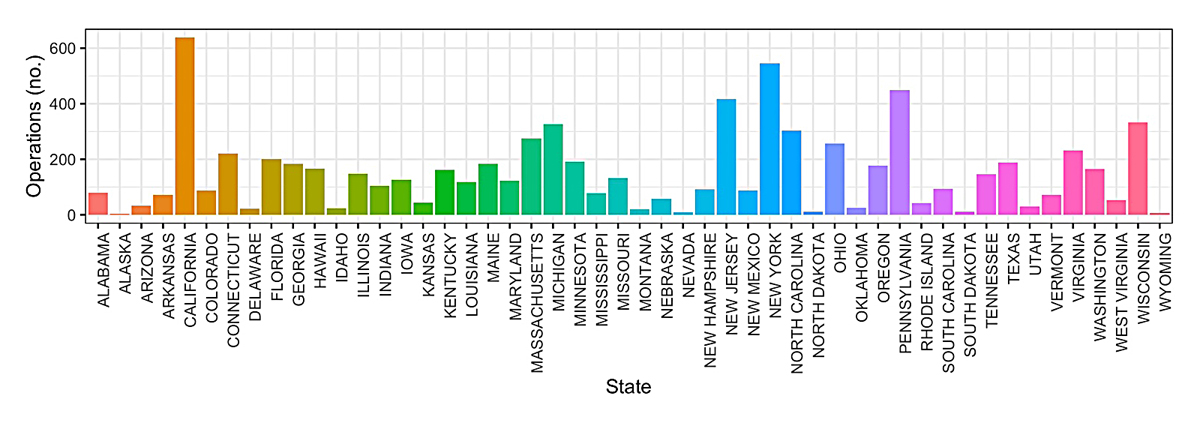 Figure 4. Total number of operations producing eggplants per U.S. state in 2017.
Figure 4. Total number of operations producing eggplants per U.S. state in 2017.
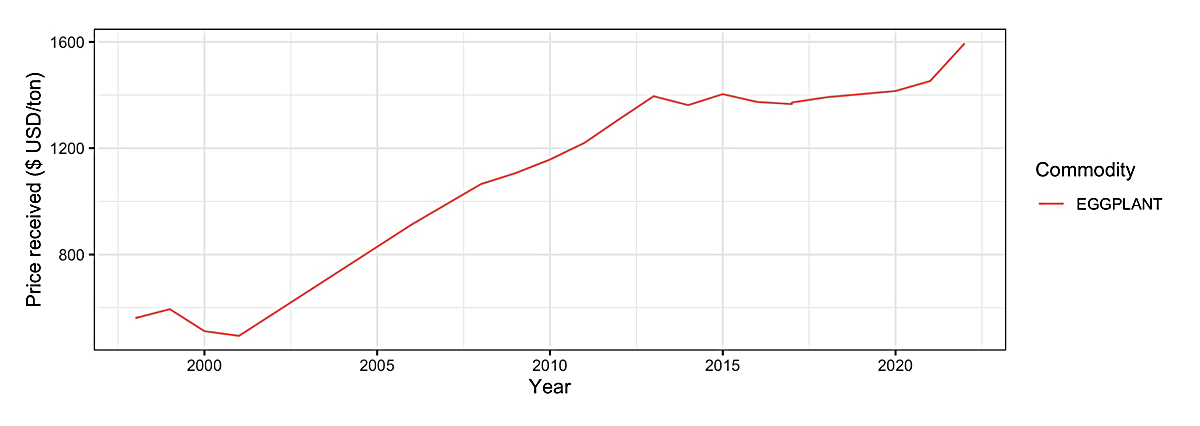 Figure 5. Price received by U.S. producers for eggplants in USD per ton (1998–2022).
Figure 5. Price received by U.S. producers for eggplants in USD per ton (1998–2022).
Among local landraces and wild relatives, eggplant displays extremely diverse variation in growth patterns, biotic and abiotic tolerance, and fruit and leaf shapes [3,6,7]. In its primitive form, eggplant is a tall, woody perennial plant that bears small, thick-skinned fruits that are bitter and seedy when mature [2,4,8,9]. The culmination of domestication, cultivation, and breeding lead to a development of smaller plant that is cultivated as an annual crop all over the world. Most cultivated varieties lack prickles and yield perfect, frequently solitary flowers. Fruits are larger, less seedy, and less bitter than wild varieties. Cultivated eggplant fruit also typically have thin skin and tender flesh. Diverse fruit morphologies, including rounded, ovate, oblong, fasciated, elongate, and serpentine, can be found among varieties. Fruit size varies widely along with shape. Fruit sizes range from 2 to 35 cm in diameter and 4 to 45 cm in length. For fruit mass, there is a 100-fold variation amongst varieties [6]. Fruit colors are attributable to the varying amounts of anthocyanin (red and purple) and chlorophyll (green) pigments in the growing fruit. A variety of colors, some with contrasting stripes or streaks have been selected for, including white, green, violet, purple, and nearly black types [7,10].
The primary goals of eggplant breeders have been to improve productivity, enhance traits related to fruit quality such as the flavor and texture of the flesh, the thickness, color, and glossiness of the skin, the capability to store the harvested fruit, size attributes of the fruit (length and girth), and incorporation of pest and disease resistance into the crop. Since heterosis in eggplant was discovered, breeding efforts have increasingly focused on producing hybrids from inbred stock that are more productive. F1 hybrids now occupy the bulk of commercial cultivars [11]. However, hybrid seed production is labor-intensive, which hampers eggplant breeding. Manually emasculating and pollinating the inbred parents involves a significant amount of time and money. As a result, attempts are being made to introduce CMS into eggplant breeding lines [12,13].
Recently, breeding for parthenocarpy has drawn considerable attention (the development of seedless fruits). Under otherwise unfavorable climatic circumstances, like sub or supra optimal temperatures and humidity levels, low light, heavy rain, or high winds, parthenocarpy ensures fruit establishment and growth. Additionally, compared to their seeded counterparts, seedless fruits often have flesh that is softer, less bitter, and browns more slowly [14]. Another technique used to produce interspecific eggplant hybrids is somatic hybridization [15–18]. These hybrids typically display desirable phenotypes, but they also have a propensity for sterility. The tetraploid form of somatic hybrids makes it more difficult to incorporate them into breeding operations. However, another culture has shown to be effective for reverting such lines to diploidy [19,20].
The two most significant crops in the Solanaceae family, tomato (S. lycopersicum) and potato (S. tuberosum), were the primary targets of early mapping studies [21]. With the development of comparative genomics and molecular linkage maps, the molecular focus that was previously confined to tomatoes has expanded to include eggplant (as well as peppers). Based on a population of 168 F2 individuals produced from a hybrid between S. melongena breeding lines from Japan (EPL1) and India (WCGR112-8), the first linkage map for eggplant was constructed [22]. In 2014, the first draft of the eggplant genome was made public, indicating a genome size of approximately 1.13 Gb [23]. Current references indicate a genome size of between ~1.10 and ~1.25 Gb for the diploid eggplant, with a base chromosome number of 12 [3,24–26].
The first high-quality chromosome-anchored reference genome, with an estimated genome size of 1.21 Gb, and the first genome resequencing study were recently made available for the eggplant-breeding community [24,27]. The relatively late sequencing of the eggplant genome has contributed to a delay in the understanding of the genetics and genomics of relevant agronomic traits, which so far has been limited to genome-wide association study (GWAS) and mapping in biparental populations. However, substantial work has been done in recent years to create novel germplasm materials that will aid in the analysis of complex and quantitative traits, with many of these attempts exploiting wild relatives of eggplant as a source of variation [28–33]. Doganlar et al. [34] and Frary et al. [35] investigated the genetic bases underlying a wide range of complex morphological characters of eggplant using the S. linnaeanum MM195 × S. melongena MM738 F2 mapping population. Fourteen domestication-related characteristics, including fruit shape and size, prickliness, and color of leaves, stems, and fruit, were measured in two locations [34]. At both sites, 15 QTLs were shown to be significantly influencing corresponding traits. This led to the understanding that a few loci on linkage groups 2, 7, and 9 controlled the majority of the variation in fruit size (fw2.1 and fw9.1). and fruit shape (fs2.1 and fs7.1). These genes are more widely conserved throughout Solanaceae family, as shown by the mapping of the identical QTLs in tomato [36,37] and pepper [38]. The Solanaceae has become a model for comparative genomics due to the close kinship of eggplant, tomato, and pepper, which has also aided the creation of various molecular techniques and genetic maps in eggplant. These results also imply that the similarities in gene structure and function between the tomato and eggplant genomes should make it possible to use what is known about tomatoes to improve eggplant [6].
Frary et al. [35] found 63 significant QTLs in an assessment of 18 traits that influence leaf, flower, and fruit size, shape, appearance, and development. These traits have major impacts on fruit yield and quality; thus, wild germplasm is a valuable genetic resource for the improvement of complex agronomic traits in cultivated eggplant.
One of the most crucial attributes in the characterization and breeding of eggplants is fruit morphology [39]. In fact, fruit shape is the primary factor used to categorize eggplant cultivars [39,40]. Commercially, there are three main types of eggplant cultivars: Round, semi-long, and long. [40–42]. In eggplant, even within varietal groups, there is a great deal of variation in fruit shape (Figure 6) [39–41,43,44], much as there is with other Solanaceae crops domesticated for their fruits, like tomato or pepper [38,45–47].
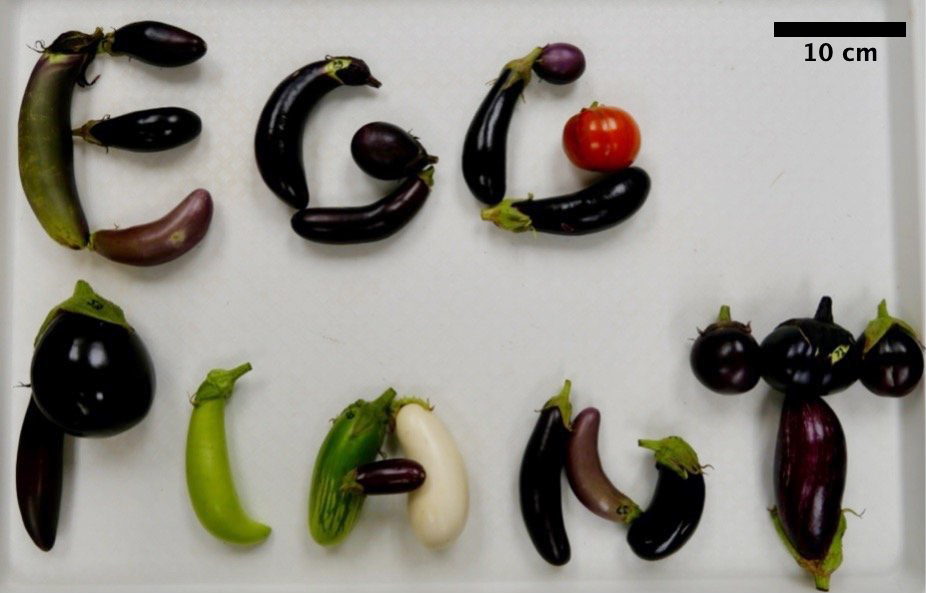 Figure 6. Fruit shape, size, and color variation within cultivated eggplants. *Note: orange fruit used in the second G of “Eggplant” belongs to species S. aethiopicum (also known as Ethiopian eggplant, bitter tomato, nakati, pumpkin-on-a-stick, and mock tomato) all other fruit are representative of variation within S. melongena, eggplant.
Figure 6. Fruit shape, size, and color variation within cultivated eggplants. *Note: orange fruit used in the second G of “Eggplant” belongs to species S. aethiopicum (also known as Ethiopian eggplant, bitter tomato, nakati, pumpkin-on-a-stick, and mock tomato) all other fruit are representative of variation within S. melongena, eggplant.
For breeders, minor variations in eggplant fruit morphology are important and may determine the success or failure of a commercial cultivar. Numerous analyses of the inheritance of fruit shape in eggplant have led to the identification of several QTLs for this fruit characteristic [34,40]. Phenomics tools that offer a precise and thorough description of fruit shape are needed for further advancements in this sector of eggplant research. Studies on eggplant fruit morphology characterization typically rely on straightforward traits measured manually, such as fruit length or width [34,40,41]. Most studies examined in this paper measured these traits manually. Fruit width was typically measured at the fruit's widest point, while fruit length was measured from the fruit’s base to its tip. The height of the plant is generally determined by utilizing a meter ruler to measure the distance between the soil’s surface and the main stem’s apical point at flowering. The number of days to first flowering is observed as the number of days from transplanting to the anthesis of first flower following the common protocols for manual characterization of eggplant. Even though these traits are helpful and easier for non-specialized staff to measure, they may not capture a precise characterization of fruit shape, which may be necessary for a variety of objectives, including the thorough description of germplasm accessions and cultivars, as well as selection and breeding [48]. Recently, a free program called “Tomato Analyzer” was created to analyze the tomato fruit’s shape which can also be applied to eggplant [46,47,49,50]. Tomato Analyzer facilitates fruit shape phenomics investigations since it can evaluate a wide range of fruit shape characteristics from scanned images of fruit sections. But only a couple studies included in the current analysis were used tomato analyzer software to measure fruit characteristics. The morphological characterizations for eggplant have also been defined by the European Cooperative Programme for Plant Genetic Resources [51,52]. This provides a useful protocol to follow in characterization of eggplant traits for breeding.
The percentage of phenotypic variance among individuals in a population that results from variations in genetic values is known as heritability. In other words, heritability describes how accurate the phenotypic value is as a breeding value indicator. In this context, high heritability here refers to a high level of similarity in a particular attribute between parents and offspring, and low heritability refers to a low level of similarity [53]. The genetic make-up of the plant, the environment in which it is cultivated, and the interplay of the genotype and environment cumulatively influence plant phenotype.
A plant breeder’s objective is to select plants with genotypes that give superior phenotypes rather than plants with favorable phenotypes brought on by environmental factors. When genetic variance accounts for the majority of variation in a trait, traits have higher heritability and are generally easier to advance more swiftly through selection. Heritability is also widely used by plant breeders to calculate the precision of a single field experiment or a set of field trials [54]. The reliability of phenotypic value as an indicator of a plant’s breeding value can also be expressed through heritability. Practically speaking, breeders may only assess an individual’s phenotypic value; however, the breeding value must be estimated as it determines how that individual will influence the following generation. As a result, the degree to which a breeder may enhance a trait in the next generation by choosing parents based on an individual’s phenotypic value will depend on how closely the phenotypic value corresponds with the breeding value [53].
Total phenotypic variance in a population can be partitioned into genotypic, environment, and genotype × environment (G × E) interaction variances. This is equationally represented as:
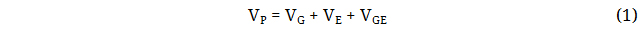
Where VP represents the total phenotypic variance, VG represents the total genotypic variance, VE denote the environmental contribution to the variance and VGE represents the interaction. The genetic component of the variation can be further divides into additive genetic variance (VA), dominance genetic variance (VD) and epistatic genetic variances (VI):
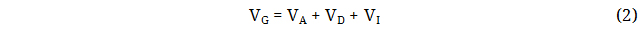
The sum of the effects of all the alleles an individual possesses is their additive genetic variance, also known as breeding value [53]. Epistatic variance comprises interactions between loci, whereas dominance variance includes interactions of alleles within a locus. Broad sense heritability (H2) is the proportion of total phenotypic variance that is due to total genetic effects that may include dominance and epistasis.
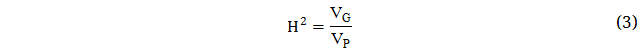
Simultaneously, the narrow sense heritability (h2) is the proportion of total phenotypic variance that is due to additive genetic effects, and thus most immediately applicable to selection. Narrow sense heritability estimates are resultingly generally lower than broad sense heritability estimates as they do not include dominance or epistatic genetic variances.
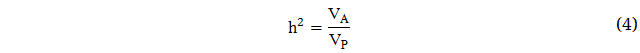
Narrow sense heritability (h2) is oftentimes more relevant in the selection process as a result of the greater dependency of response to selection on the additive effect of the alleles [55,56]. Heritability ranges between 0 and 1 given that it is defined as a ratio of variance components.
Heritability predicts the response to selection for a given trait and population. Usually, parents are selected from a base population based on superior performance. The difference between means of the selected parents (µs) to the mean of base population (µ0) is called selection differential (S = µs–µ0). The selection response (R) is the measure of how much genetic gain obtained from the population created by crossing the selected parents. Because the genetic component that results from additive genetic variance is measured by narrow sense heritability, multiplying the narrow sense heritability by the selection differential will yield the response to selection (R = h2S). Thus, the anticipated percentage of the selection differential that will be realized as a benefit from selection or heritability is (h2 = R/S).
The main goal of estimating heritability is to compare the genetic gains from selection (genetic advance) based on alternative selection strategies under different experimental designs and environments. This knowledge, along with the relative costs of further replications, locations, and years, can be used to formulate the most effective breeding schemes [57]. Estimates of heritability are unique to the population and environment for which they are computed. Therefore, the best selection technique can be determined by comparing heritability estimates corresponding to responses to selection based on means of overall environments with heritability based on means of within subset of local environments [58–60]. The population structure used to estimate variance components, the experimental design, and the environment of evaluation all affect the calculation of heritability. Changes in the genetic or environmental components of the variance can lead to changes in both narrow and broad sense heritability estimates. Similarly, heritability estimated based on different family structures derived from the same base population can be compared to determine which family structure is best for maximizing genetic gain over unit time [61].
Due to the importance of trait heritability in plant breeding and selection, this brief review assessed 60 journal articles published during the period of 2006 to 2022 (Figure 7). This information was collected to inform new and existing eggplant breeding efforts about the proportion of phenotypic variance derived from genetic factors, the broad-sense heritability, across diverse populations and phenotyping environments. Examined in this brief review are broad-sense heritability estimates obtained from studies based on different types of family structures, different screening environments, and different variety trials.
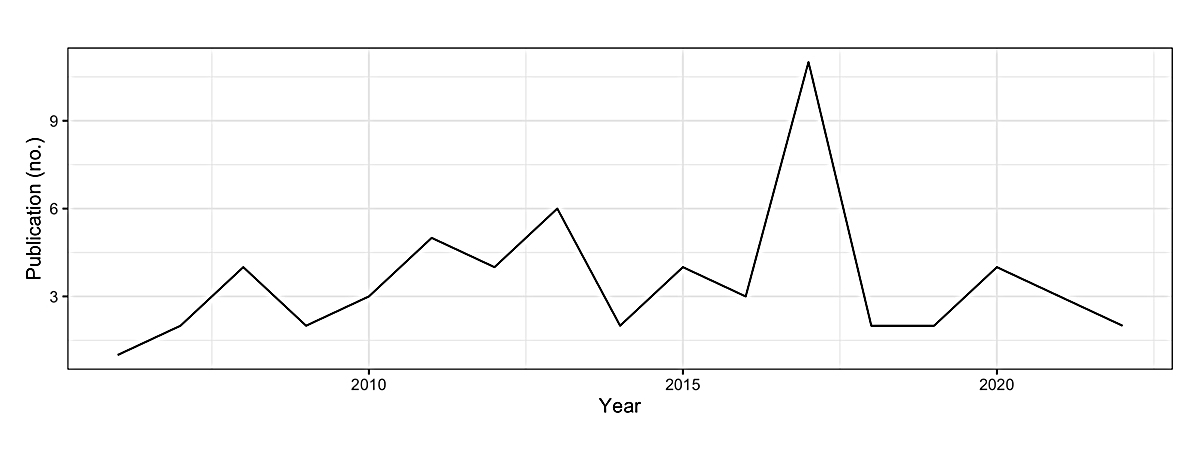 Figure 7. Yearly distribution of the number of published studies on the heritability of horticultural traits in eggplants examined in this review [62–121].
Figure 7. Yearly distribution of the number of published studies on the heritability of horticultural traits in eggplants examined in this review [62–121].
Data were compiled from published scientific articles using the CAB Direct database starting with the search terms “solanum melongena”, “heritability”, and “yield” [122]. A pre-screening of potential horticultural traits for assessment beyond yield was conducted for the first 30 papers accessed to assess the prevalence of other traits in the literature. Data were then extracted from publications for the nine additional traits that occurred with high frequency in heritability studies of eggplants with a focus on work conducted in S. melongena.
The examined traits included plant architecture (branch number [BNO] and plant height [PLHT]), phenology (days to first flower [DFFL] and days to fifty percent flowering [D50F]), and fruit size/yield related traits (fruit length [FRLN], fruit diameter [FRDM], fruit girth [FRGR], fruit number [FRNO], individual fruit mass [FRMS], and yield [YLD]). A total of 188 papers were examined, of which 60 papers met the criteria of depicting broad-sense heritability of at least one of the traits of interest. Broad-sense heritability estimates were extracted from each manuscript and standardized to a value out of 1.000 if they were depicted on a scale of 0 to 100. Where negative broad-sense heritability values or values greater than 1.000 were observed, these values were omitted from evaluation.
Broad-sense heritability estimates were based on different relationships and included Fn populations (F1, F2, F3, F4, F5, F6), back-cross populations (BC1, BC2, BC3, BC3F1, BC3F2, BC3F3), generation mean analysis structured populations, half-diallel populations, North Carolina Design I, introgression lines, mass selection populations, line by tester populations, and wide evaluations of germplasm, cultivars, and local accessions.
Across the traits examined, all ten were observed to have a maximum broad-sense heritability estimated above 0.990 (Table 1). The lowest overall minimum observed broad-sense heritability estimates were for traits such as plant height, branch number, days to first flower, fruit length, single fruit mass, and yield, which fell below 0.100 for the lowest observed values. Six of the lowest observed heritability estimates for plant height came from the same study which examined F2, BC1, and BC2 progenies. The total number of studies reporting broad-sense heritability estimates for traits ranged from 60 (plant height) to 24 (days to fifty percent flowering). Many studies reported multiple broad-sense heritability traits based on different populations or environments evaluated, as a result four traits (plant height, fruit number, fruit length, and yield) all had over 100 cumulative broad-sense heritability estimates.
The mean broad-sense heritability estimates for the ten evaluated traits ranged from 0.661 (branch number) to 0.800 (fruit diameter) when evaluating the mathematical mean across all observations (Table 2). For the mathematical mean weighted by study, such that only the mean of each study was assessed, values ranged from 0.695 (branch number) to 0.845 (individual fruit mass). Beyond branch number, all traits exceeded 0.700 for broad-sense heritability when weighted by study. Broad-sense heritability of fruit number (0.837), fruit length (0.818), fruit diameter (0.800), and individual fruit mass all where at or above 0.800 when weighted by study, while only fruit diameter (0.800) was at that level when examining the mean across all observations.
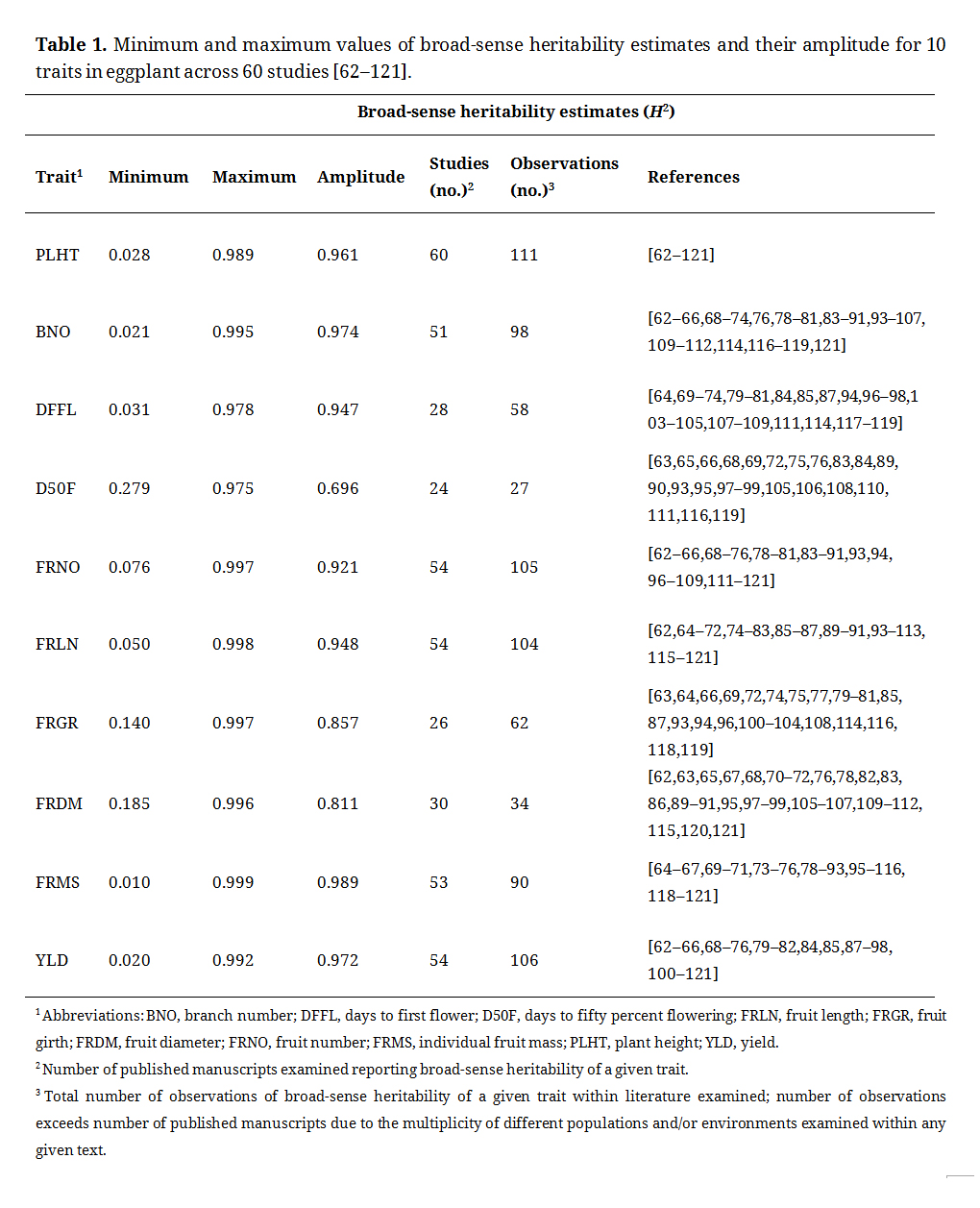 Table 1. Minimum and maximum values of broad-sense heritability estimates and their amplitude for 10 traits in eggplant across 60 studies [62–121].
Table 1. Minimum and maximum values of broad-sense heritability estimates and their amplitude for 10 traits in eggplant across 60 studies [62–121].
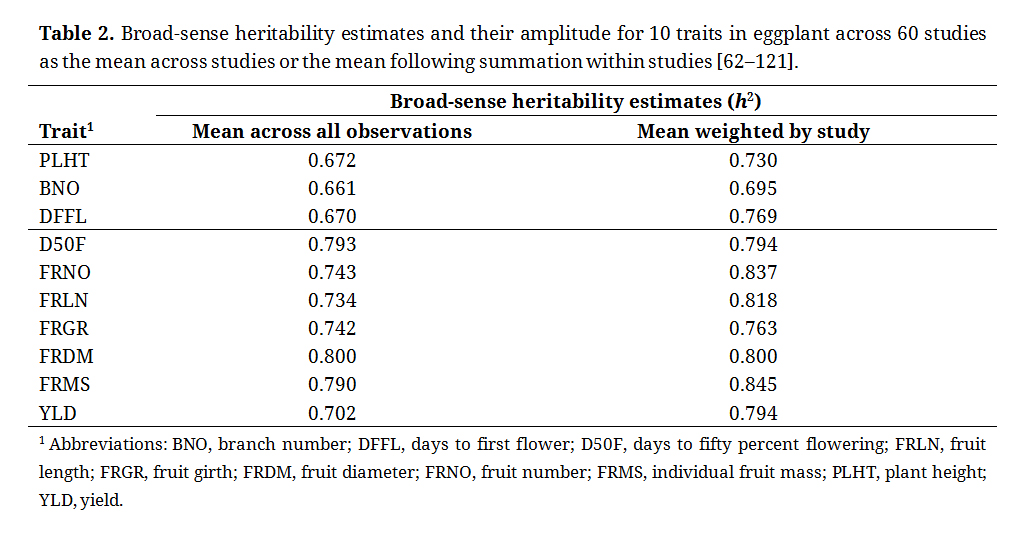 Table 2. Broad-sense heritability estimates and their amplitude for 10 traits in eggplant across 60 studies as the mean across studies or the mean following summation within studies [62–121].
Table 2. Broad-sense heritability estimates and their amplitude for 10 traits in eggplant across 60 studies as the mean across studies or the mean following summation within studies [62–121].
Traits that have lower heritability may be more difficult to select for regardless of environmental conditions or population architecture. Additionally, the genetic component of traits with lower heritability may be more complex, involving multiple genes and interactions, which can make it challenging to identify the specific genetic factor that are responsible for the trait. Bias may come from estimating heritability in germplasm evaluations or variety trials rather than in on-going breeding populations. Studies using germplasm evaluations and variety trials typically involve a small sample of varieties or genotypes, which may not be representative of the entire population of genetic variability. This sample bias can lead to an overestimate of heritability if the sample has a higher proportion of genetic variation than the population. Germplasm evaluations typically contains individuals from diverse backgrounds. This diversity also can lead to bias in the heritability estimate because the genetic variation among individuals is not solely due to the genetic factors that are being evaluated for the trait. In contrast, heritability estimates from breeding populations are typically more accurate because the individuals being evaluated are more genetically similar. This is because breeding populations are typically created by crossing individuals that have similar genetic backgrounds, resulting in a more homogeneous population with less variation due to environmental factors. To overcome some of these problems, it’s important to have a good experimental design, a large sample size, a controlled environment, and a precise measurement of the trait of interest.
Further examination of heritability was conducted via partitioning of the broad-sense heritability estimates into two main pools as (1) heritability calculated based on genotype performance in a wide variety trial and (2) heritability calculations based on specific mating designs and interrelationships among individual seedlings. For plant architecture and phenological traits (Figure 8), broad-sense heritability estimates were always greater for the calculations based on genotype performance. When accounting for mating design, all four of these traits had mean broad-sense heritability estimates below 0.650.
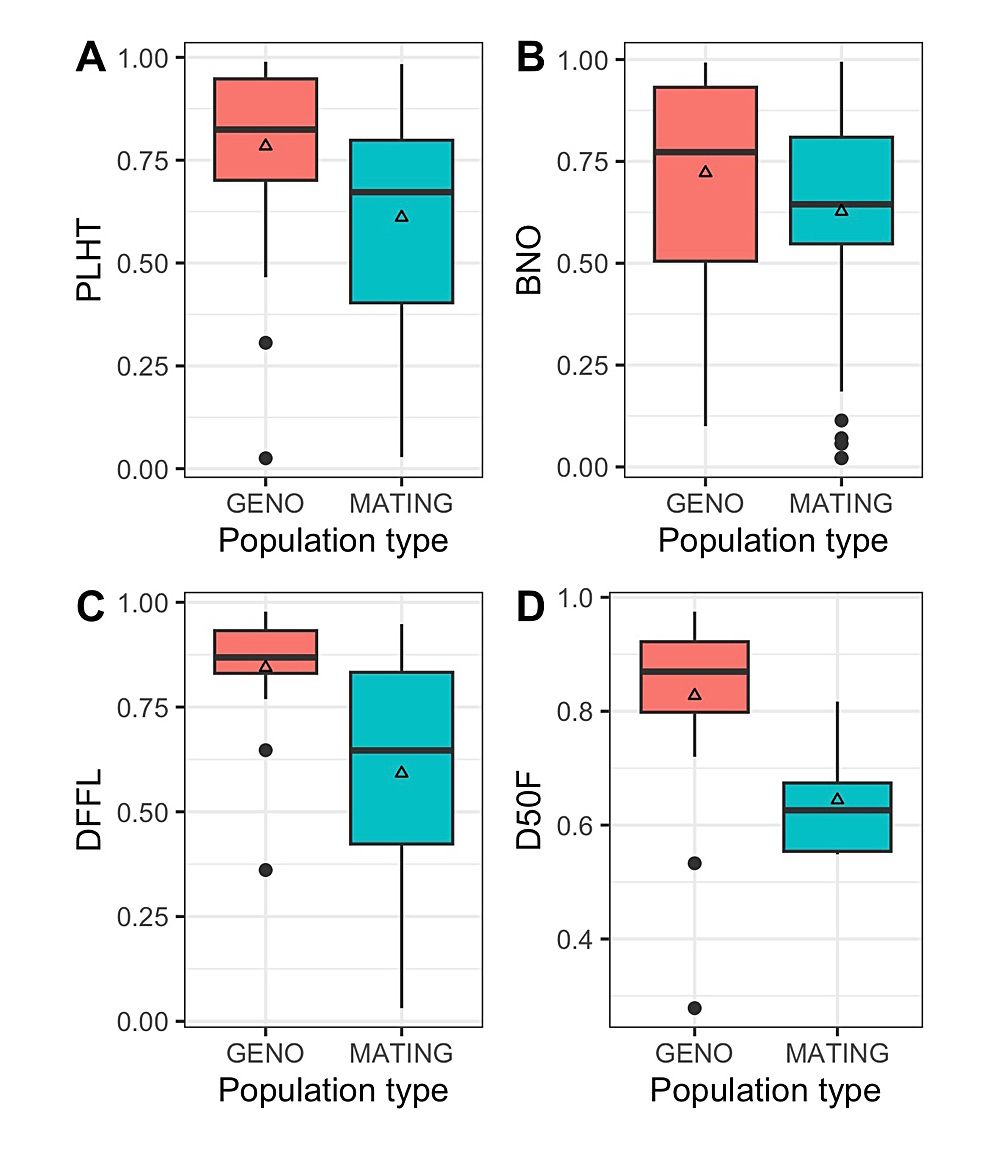 Figure 8. Boxplots showing the distribution broad-sense heritability estimates of two plant architectural traits, (A) PLHT = plant height and (B) BNO = branch number, and two phenological traits, (C) DFFL = days to first flowering and (D) D50F = days to fifty percent flowering, based on population type under examination (GENO = broad-sense heritability calculated with germplasm screening/variety trial data from distinct genotypes; MATING = heritability estimates calculated based on different structure populations in which individuals have defined genetic relationships). Note: Open triangle indicates the mean broad-sense heritability, black line indicates median broad-sense heritability, upper and lower portions of boxplot indicate upper and lower quartile, whiskers indicate upper (largest) and lower (smallest) value ranges that are not outliers, filled black circles indicate outlier values.
Figure 8. Boxplots showing the distribution broad-sense heritability estimates of two plant architectural traits, (A) PLHT = plant height and (B) BNO = branch number, and two phenological traits, (C) DFFL = days to first flowering and (D) D50F = days to fifty percent flowering, based on population type under examination (GENO = broad-sense heritability calculated with germplasm screening/variety trial data from distinct genotypes; MATING = heritability estimates calculated based on different structure populations in which individuals have defined genetic relationships). Note: Open triangle indicates the mean broad-sense heritability, black line indicates median broad-sense heritability, upper and lower portions of boxplot indicate upper and lower quartile, whiskers indicate upper (largest) and lower (smallest) value ranges that are not outliers, filled black circles indicate outlier values.
For fruit related traits, the same trend was observed, with greater broad-sense heritability estimates based on performance of individual genotypes (Figure 9). Mean broad-sense heritability estimates ranged from 0.841 (fruit diameter) to 0.896 (fruit girth) based on performance of individual genotypes. Contrastingly, mean broad-sense heritability estimates ranged from 0.618 (yield) to 0.726 (individual fruit mass) when calculated based on studies using mating designs in heritability estimates.
Broad-sense heritability estimates of fruit shape (fruit length, diameter, and girth) were generally high based on genotype performance (>0.850). Fruit shape may be readily selected for in these types of populations. However, for mating populations, although broad-sense heritability exceeded 0.650 for all three fruit shape traits, selection may be improved through increased phenotypic accuracy. This may be accomplished through increasing plot number, sample number, or sample throughput through image-analysis pipelines capable of capturing fruit shape measurements, such as Tomato Analyzer software [46,47]. For fruit number (0.672), single fruit mass (0.726), and yield (0.618), improvements may also be necessary to increase genetic gain from selection each cycle considering the lower levels of heritability observed in structured mating studies for these three yield components.
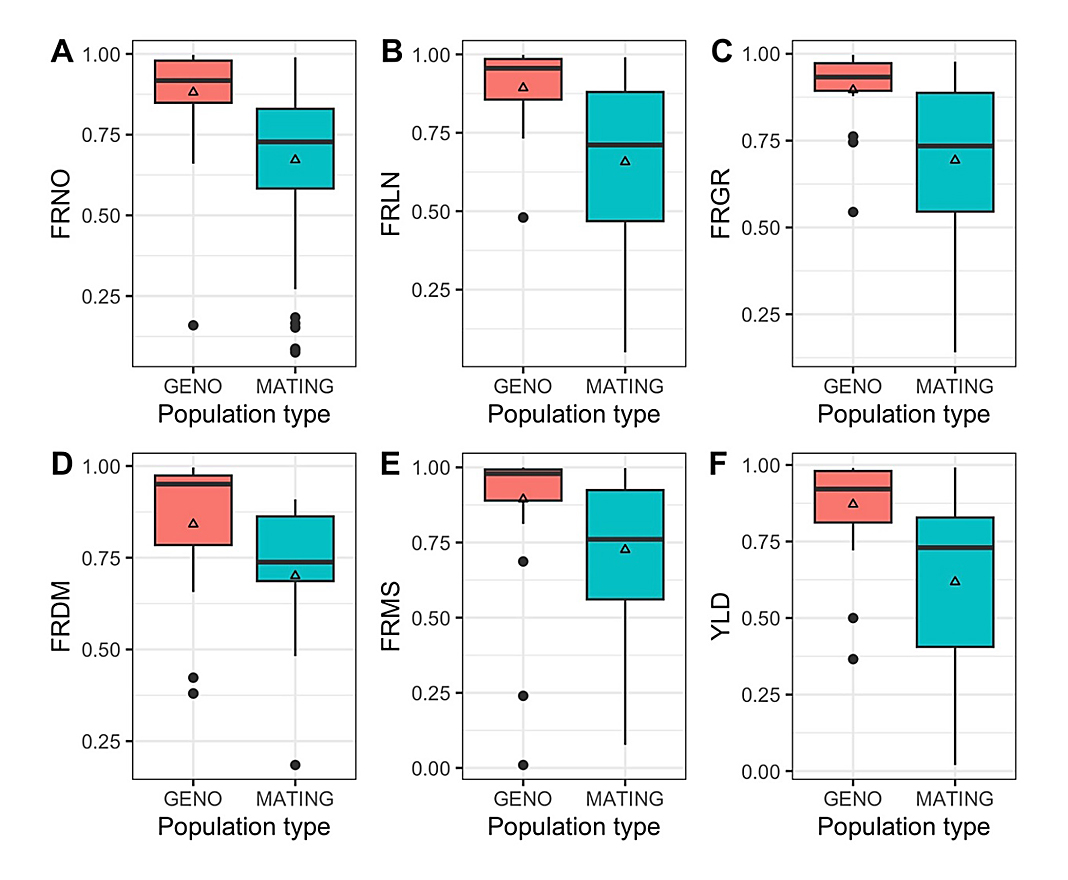 Figure 9. Boxplots showing the distribution broad-sense heritability estimates of six fruit related traits, (A) FRNO = fruit number, (B) FRLN = fruit length, (C) FRGR = fruit girth, (D) FRDM = fruit diameter, (E) FRMS = single fruit mass, and (F) YLD = yield, based on population type under examination (GENO = broad-sense heritability calculated with germplasm screening/variety trial data from distinct genotypes; MATING = heritability estimates calculated based on different structure populations in which individuals have defined genetic relationships). Note: Open triangle indicates the mean broad-sense heritability, black line indicates median broad-sense heritability, upper and lower portions of boxplot indicate upper and lower quartile, whiskers indicate upper (largest) and lower (smallest) value ranges that are not outliers, filled black circles indicate outlier values.
Figure 9. Boxplots showing the distribution broad-sense heritability estimates of six fruit related traits, (A) FRNO = fruit number, (B) FRLN = fruit length, (C) FRGR = fruit girth, (D) FRDM = fruit diameter, (E) FRMS = single fruit mass, and (F) YLD = yield, based on population type under examination (GENO = broad-sense heritability calculated with germplasm screening/variety trial data from distinct genotypes; MATING = heritability estimates calculated based on different structure populations in which individuals have defined genetic relationships). Note: Open triangle indicates the mean broad-sense heritability, black line indicates median broad-sense heritability, upper and lower portions of boxplot indicate upper and lower quartile, whiskers indicate upper (largest) and lower (smallest) value ranges that are not outliers, filled black circles indicate outlier values.
Broad-sense heritability estimates for horticultural traits of eggplant vary based on the population in which traits were measured. Higher estimates of broad-sense heritability were frequently observed when estimated in germplasm or variety screening trials compared to breeding populations. Estimates of broad-sense heritability exceeded 0.500 for all traits. The yield components of single fruit mass and fruit number per plant had greater heritability estimates than yield itself. Phenology and plant architecture traits’ broad-sense heritability were lower than yield components. Broad-sense heritability estimates for all traits indicate selection effectively garner a response. Where heritability estimates are lower, methods can be improved to reduce non-genetic variance (improve measurement accuracy through incorporating image analysis or other quantitative approaches).
The dataset of the study is available from the authors upon reasonable request.
VK and AS designed the review, VK collected the metadata, VK and AS analyzed the data, VK and AS created data visualizations, and VK and AS developed the manuscript.
The authors declare that they have no conflicts of interest.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60.
61.
62.
63.
64.
65.
66.
67.
68.
69.
70.
71.
72.
73.
74.
75.
76.
77.
78.
79.
80.
81.
82.
83.
84.
85.
86.
87.
88.
89.
90.
91.
92.
93.
94.
95.
96.
97.
98.
99.
100.
101.
102.
103.
104.
105.
106.
107.
108.
109.
110.
111.
112.
113.
114.
115.
116.
117.
118.
119.
120.
121.
122.
Kadium VR, Svyantek A. Broad-Sense Heritability for Horticultural Production Traits in Eggplant. Crop Breed Genet Genom. 2023;5(2):e230003. https://doi.org/10.20900/cbgg20230003

Copyright © 2023 Hapres Co., Ltd. Privacy Policy | Terms and Conditions




